Реплікація включає: ініціацію, елонгацію та термінацію.
Ініціація реплікації.Цей процес включає утворення реплікативної вилки та синтезу праймера.
1. Утворення реплікативної вилки
Оскільки для забезпечення напівконсервативного механізму реплікації ДНК необхідна наявність одноланцюгової матриці, важливою умовою початку ініціації є дестабілізація структури ДНК та розкручування хеліксу з утворенням реплікативної вилки.
У бактерії Е. соlі реплікація розпочинається зі специфічного сайту початку реплікації – точки ori С, яка містить специфічні чергування нуклеотидів, що зв’язують білкові фактори (Dna В, Dna С, Dna G). Ці білкові фактори забезпечують розкручування хеліксу ДНК та синтез праймера. Для них характерна хеліказна, топоізомеразна, гіразна, АТФ-азна та праймазна активність.
Етапи утворення реплікативної вилки:
- дестабілізація суперспіралізованої структури ДНК,
- розкручування двоспіральної структури (хеліксу) ДНК,
- руйнування водневих зв’язків між комплементарними парами азотистих основ дезоксирибополінуклеотидних ланцюгів молекули ДНК,
- розходження ланцюгів ДНК з утворенням Y-подібної реплікативної вилки.
2. Синтез праймера
Праймери мають РНК-ову структуру, оскільки відбір рибонуклеозидтрифосфатів із середовища відбувається за участю ДНК-залежної РНК-полімерази (праймази). Праймери являють собою олігонуклеотидні фрагменти, які включають 8-10 рибонуклеотидів і містять на 3/ кінці вільну ОН-групу.
Синтез праймерів каталізує мультиферментний комплекс – праймосома, яка є компонентом складнішого комплексу – реплісоми, до складу якого входить цілий ряд ферментів та білкових факторів.
Розпочинається синтез праймера із приєднання ДНК-зв'язуючого білка до стартової точки ведучого дезоксирибополінуклеотидного ланцюга матриці. Далі, за участю праймази, проходить синтез затравного ланцюга РНК-ової структури, комплементарного до матриці ДНК.
Після закінчення синтезу на ведучому ланцюгу, праймосома приєднується до ланцюга, що запізнюється та за участю β-білка ініціює синтез на ньому не одного, а кількох праймерів.
Праймосома рухається по цьому ланцюгу від реплікативної вилки, протилежно до ведучого ланцюга ДНК, однак в напрямку 3/ → 5/.
Тобто синтез праймера (затравного ланцюга з РНК-ковою структурою) проходить майже одночасно як на ведучому, так і на запізнюючому ланцюгах.
Для кільцевих молекул ДНК прокаріот характерна двоспрямована реплікація: реплікативні вилки від точки ori рухаються у двох напрямках:
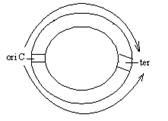
За участю хеліказної та праймазної активності білкових факторів проходить їх спряжене функціонування: хелікази розплітають дуплекс ДНК, а праймази забезпечують синтез праймерів.
Необхідність такого механізму ініціації зумовлена тим, що ДНК-полімераза III не може самостійно розпочати синтез ДНК на структурі матриці з використанням вільних нуклеозид-5/-трифосфатів. Тому для початку синтезу ДНК (ініціації дії ДНК-полімерази ІІІ) необхідна затравка, яка б містила вільну 3'-ОН-групу, за допомогою якої може відбуватися полімеразна реакція – подовження полінуклеотидного ланцюга ДНК (“ріст з хвоста”).
Елонгація реплікації включає процеси:
- подовження полінуклеотидних ланцюгів ДНК;
- сполучення синтезованих фрагментів з утворенням комплементарних копій (дочірніх ланцюгів).
1. Подовження полінуклеотидних ланцюгів ДНК.
За участю ДНК-залежної ДНК-полімерази ІІІ, для якої характерна ДНК-полімеразна та нуклеотидилтрансферазна активність, відбувається нарощування дезоксирибополінуклеотидного ланцюга.
ДНК-залежна ДНК-полімераза III одночасно активує два субстрати:
- з’єднаний за принципом комплементарності з матрицею кінець затравки, яка містить вільну З'-ОН-групу рибози,
- дезоксирибонуклеозид-5-трифосфати (дНТФ), що надходять з середовища.
За участю ферменту проходить нуклеофільна атака α-фосфату дезоксирибонуклеозидтрифосфату 3'-ОН-групою, яка міститься на кінці затравки, внаслідок чого дезоксирибонуклеозид-5/-монофосфат приєднується до атома Оксигену гідроксильної групи в 3/-положенні. Результатом цього є утворення 5/→3/-фосфодиефірного зв'язку та вивільнення пірофосфату. Продукт реакції містить вільну 3'-ОН-групу, по якій знову повторюється полімеразна реакція, що дістала назву „ріст з хвоста”:
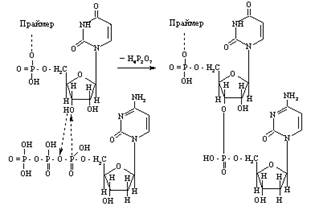
У результаті багаторазового повторення цієї реакції відбувається поступове комплементарне нарощування дочірнього ланцюга.
Реплікація запізнюючого ланцюга відбувається фрагментарно, оскільки на цьому ланцюгу ініціюється синтез не одного праймера, а їх великої кількості, внаслідок подовження яких по 3'-ОН-групи утворюються фрагменти, які дістали назву “фрагментів Оказакі”. Чергування дезоксирибонуклеотидів на синтезованих фрагментах комплементарні до відповідної ділянки матричного ланцюга.
Поступове нарощування фрагментів Оказакі ферментом ДНК-залежною ДНК-полімеразою триває доки не досягне місця локалізації наступної затравки, з якої розпочинається ріст нового фрагменту. Ведучий і запізнюючий ланцюги ДНК реплікуються синхронно завдяки участі в цьому процесі димерного ДНК-полімеразного комплексу:
2. Сполучення синтезованих фрагментів з утворенням комплементарних копій (дочірніх ланцюгів)
Для з’єднання фрагментів Оказакі необхідне видалення праймерів з РНК-овою структурою. Це відбувається внаслідок дії специфічної нуклеази (Н-РНК-ази), або 5/→3/-екзонуклеазної активності ДНК-полімерази І.
Після видалення рибонуклеотидів праймера проходить їх заміна на відповідні дезоксирибонуклеотиди. Далі, кінці прогалин, за участю ферменту ДНК-лігази, з’єднуються між собою, внаслідок чого формується суцільний запізнюючий дочірній ланцюг ДНК.
Репліковані ланцюги утворюють дві дочірні молекули ДНК, кожна з яких містить один батьківський та один дочірній ланцюги і є точною копією вихідної молекули.
Термінація реплікації. Механізм термінації реплікації, або завершення синтезу ДНК, має свої особливості. В геномі деяких бактерій термінація відбувається за участю специфічного термінатора, в інших геномах такий термінатор відсутній. Тому припускають, що реплікація у прокаріот завершується після того, як вичерпується матриця (у бактерій, які містять лінійні дволанцюгові ДНК) або коли при подвоєнні кільцевих молекул ДНК зустрічаються дві реплікативні вилки. Закінчення процесу реплікації у клітинах бактерії Е. соlі відбувається на сайті ter, розміщеному на протилежній до точки ori ділянці кільцевої молекули.
Реплікація проходить з високою точністю та швидкістю: 1-2 тис. нуклеотидів за секунду у прокаріот і близько 100 нуклеотидів у еукаріот.
Дата добавления: 2022-05-27; просмотров: 75;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
