Экологическая генетика лесных древесных пород
Общие положения и определения
Экологическая генетика как самостоятельное направление научных исследований, синтезирующее достижения в области генетики и экологии, возникла в связи с все возрастающим загрязнением биосферы, интенсификацией промышленности, сельскохозяйственной, лесохозяйственной и других видов деятельности, увеличением энергозатрат на производство единицы продукта, снижением устойчивости новых высокопродуктивных сорго к неблагоприятным воздействиям окружающей среды.
Экологическая генетика тесно связана с популяционной генетикой,количественной генетикой, популяционной биологией и экологией. Одни лесные специалисты (Heslop-Harrison, 1964 — цит. по Р. М. A. Tigerstedt, 974) считают, что возникновение этой науки связано с началом изучения экотипов лесных древесных пород, т. е. примерно с 20-х годов XX века, обосновывают это тем, что учение об экотипах включает идеи и методы генетики, таксономии, физиологии растений и экологии. Другие идут еще дальше. Так, О. Langlet, 1971 (цит. по Р. М. A. Tigerstedt, 1974) считает, что генетико-экологические исследования насчитывают 200 лет и связаны исследованием значения географического происхождления семян в лесном деле.
Не вдаваясь в детали дискуссии, следует отметить, что, во-первых, генетическая экология как современная наука первоначально большее распространение получила при исследовании животных; во-вторых, генетическая экология является более широкой областью науки, чем исследования по экотипам; и в-третьих, учение об экотипах, конечно же, является существенной основой для генетико-экологических исследований.
Предметом исследований экологической генетики являются адаптивные реакции организмов, особенности их модификационной и генотипической изменчивости в системе “генотип — среда”. И на это особое внимание обращают такие отечественные ученые, как С.С.Шварц (1974), . А. Жученко (1980, 1995), И. А. Захаров (1984), В. А. Драгавцев (1997).
В последнее время подход к определению предмета экологической генетики расширился. В частности, С. Г. Инге-Вечтомов (1999) выделяет следующие основные разделы экологической генетики: симбиогенетика, устойчивость организмов к факторам окружающей среды, мутагенез, генетическая токсикология, прогнозирование и поиски путей предотвращения эпидемий и др. Для лесных древесных растений наибольший интерес представляют два первых направления.
Симбиогенетика охватывает целый ряд проблем современной биологии: от симбиогенеза, характерного для системы эукариотической клетки, совмещающей черты прокариотической (митохондрии, пластиды и другие клеточные органеллы) и истинно эукариотической (цитоплазма) организации, до симбиотических отношений растений и почвенных микроорганизмов. Во взаимодействии сельскохозяйственных растений и клубеньковых бактерий выявлена целая система генов, управляющих симбиотическими отношениями как со стороны бактерий, так и со стороны растений. В последние годы было установлено, что взаимодействие бобовых растений и клубеньковых бактерий осуществляется через специальную генетическую систему.
Идентифицированы первые растительные морфогены, индуцирующие развитие симбиотических органов — корневых клубеньков. С другой стороны, растения-хозяева, на которых индивидуальные штаммы ризобий (микориз, бактерий) формируют клубеньки, детерминируются бактериальными Nod-генами (nodule с англ. — узелок, клубенек). Некоторые из этих генов кодируют белки определяющих биосинтез сигнальных молекул (Nod-факторов). Комплементарность взаимодействия между штаммами ризобий и видами (сортами) бобовых в значительной степени определяется структурным разнообразием Nod-факторов, которое, в свою очередь, определяет специфичность взаимодействий бактерий с бобовыми растениями (И. А. Тихонович,1999,2000).
Взаимодействие лесных древесных растений с различными низшими симбионтами (бактериями и грибами) на организменном уровне будет рассмотрено ниже (подраздел 4.4 и раздел 5).
Устойчивость организмов к окружающей среде и мутагенез частично уже рассматривались. Более подробно об отношении лесных древесных пород к воздействиям различных факторов окружающей среды будет изложено в последующих разделах настоящей главы.
Из вышесказанного можно сделать вывод, что экологическая генетика— это наука, изучающая наследственно обусловленные ответные реакции организмов на абиотические (в том числе антропогенные) и биотические (включающие взаимоотношения между организмами родственных и неродственных групп) факторы внешней среды. В задачи экологической генетики входит изучение механизмов ответных реакций организмов на воздействие определенных факторов внешней среды в отдельности или в их совокупности.
Таким образом, можно отметить, что экологическая генетика имеет дело с адаптацией, или приспособленностью организмов. Рассмотрим более подробно это понятие и производные от него. Под адаптацией понимается процесс или результат процесса любых изменений в структуре или функциях организма, обеспечивающих способность к существованию в данной среде. Т. Dobzhansky, 1956 и позже (цит. Р. М. A. Tigerstedt, 1974) различает понятия “адаптация” и “адаптивность”. Адаптация — это способность организмов жить и размножаться в данных условиях среды. Адаптивность — это способность к адаптации (приспособлению) в определенных границах условий внешней среды.
Те структурные или функциональные изменения организма, которые увеличивают его жизнеспособность, выживаемость, темп размножения, называются адаптивными признаками. Разная степень адаптивности, в основе которой лежит действие и взаимодействие всего генетического потенциала организма, определяет в конечном счете разную возможность генотипов приспособиться в процессе их индивидуального развития к варьирующим условиям окружающей среды.
На протяжении всей своей эволюционной жизни организмы приобретают определенную норму реакции на изменение условий окружающей среды. В связи с этим различают адаптацию генотипическую и модификационную.
Генотипическая адаптация — это адаптация, при которой изменение генотипа обеспечивает образование новой нормы реакции и гармоничное приспособление индивида к новым экологическим условиям.
Модификационная адаптация — это адаптация, позволяющая организму в пределах сложившейся нормы реакции оставаться жизнеспособным и продуцировать потомство в новых условиях среды.
Кроме того, различают онтогенетическую и филогенетическую адаптации организмов. Онтогенетическая адаптация характеризует приспособительные изменения в период индивидуального развития организма. Она обеспечивается за счет модификационной изменчивости и представляет собой динамический процесс функционально-метаболической перестройки растения. За счет онтогенетической адаптации компенсируются кратковременные изменения условий внешней среды.
Филогенетическая адаптация — это результат естественного отбора в ряде поколений. Она обеспечивает приспособленность организмов к долговременным изменениям условий внешней среды, и в основе ее лежит образование новой нормы реакции (т. е. нового генотипа).
Наличие или возникновение признаков, позволяющих организму сразу адаптироваться к условиям среды, в которых он ранее не находился, обозначается терминами преадаптация или проспективная адаптация. Та или иная структура считается преадаптированной, если она способна взять на себя новую функцию без ущерба для первоначальной.
Нередко при описании адаптивных свойств отдельного генотипа или популяции используют понятия “пластичность”, “устойчивость” или “стабильность”, “гомеостаз”, “канализация”, “буферность” и др. Они могут существенно различаться по смыслу.
Пластичность организма — это его способность проявлять определенные различия в зависимости от условий среды. Стабильность организма — это его способность сохранять свои признаки в варьирующих условиях среды неизменными. Устойчивость организма—это способность его сопротивляться воздействиям неблагоприятных факторов среды. Она может обеспечиваться как его пластичностью, так и стабильностью.
В целях упорядочения многочисленных и нередко противоречивых трактовок терминов “адаптация”, “пластичность”, “стабильность”, “устойчивость” А. А. Жученко (1980) предлагает разграничить их применительно к онтогенезу и филогенезу. В онтогенезе в основном возможна лишь модификационная изменчивость, в то время как в филогенезе может происходить генотипическая изменчивость или появление новых генетических вариантов в процессе смены поколений за счет мутаций и рекомбинаций.
Термин гомеостаз означает тенденцию системы (индивида, процесса и др.) к сохранению относительной генетической сбалансированности и восстановлению ее с помощью собственных регуляторных механизмов в случае нарушения.
Взамен термина генетический гомеостаз Allard, 1964 (цит. по А. А. Жученко, 1980) предлагает использовать термин популяционная буферность. Рассматривается два способа достижения стабильности популяции в меняющихся условиях внешней среды: а) за счет индивидуальной буферности, т. е. буферности отдельного генотипа или группы растений с одним и тем же генотипом, что равнозначно термину гомеостаз индивидуального развития; б) за счет создания буферной популяции путем смешения разных генотипов, т. е. достижения популяционной буферности или генетического гомеостаза. Сорта называют буферными, если они оказываются устойчивыми, с точки зрения продуктивности, к действию широкого диапазона факторов внешней среды.
Термин канализация обозначает способность системы, несмотря на вариабельность среды, давать определенный конечный продукт (стандартный фенотип). Канализованность, или буферность против влияния внешней среды, особенно характерна для организмов дикого типа и реже — для мутантов. Канализация развития защищает индивид не только от флуктуации среды, но в определенных пределах и от случайных отклонений от нормального фенотипа.
По мнению К. Stern и Р. М. A. Tigerstedt, 1974 (цит. по А. А. Жученко, 1980), естественный отбор оказывает основное влияние на количественные признаки, которые обычно контролируются большим числом локусов. Предполагается, например, что наиболее важные адаптивные признаки контролируются 100 и более локусами. Особенно сложным оказывается генетический контроль так называемой климатической адаптации, включающей физиологические и морфологические реакции на изменение режима влажности, освещенности, температуры и др. В определенных условиях полигены могут образовывать более крупные супергены, или коадаптированные системы генов. Считается, что появившиеся такие коадаптированные системы генов обусловливают адаптацию растений к варьирующим факторам внешней среды.
Удачный адаптивный комплекс генов в основном сохраняется даже после исчезновения экологических условий, обеспечивших его появление. Так, по данным вышеупомянутых авторов (К. Stern и Р. М. A. Tigerstedt), у произрастающей на западе Канады и в США сосны скрученной —Finns contorta Dougl. семена из шишек выделяются лишь после высокотемпературного теплового воздействия, например при лесных пожарах.
В целом генетическая система растений помимо обеспечения роста, развития и воспроизводства в обычных условиях среды накапливает и вводит в действие информацию о специфических особенностях среды популяции, а также обеспечивает возможность воспринимать и накапливать такую информацию в будущем. В этом и заключается интеграция генетики и экологии организмов в единый блок специфических особенностей, изучаемых экологической генетикой.
7.2. Особенности ответных реакций растений на воздействия условий среды
Каждый организм занимает в биосфере свою экологическую нишу, или ту часть среды обитания, которая в наибольшей степени соответствует его экологическим свойствам. По G. Е. Hutchinson, 1965 (цит. по Р. М. A. Tigerstedt, 1974), ниша популяции (или вида) представлена сетью точек в абстрактном гиперобъеме с определенными координатами, каждая из которых соответствует фактору среды (температура, влажность, засоленность и т. п.). Каждая популяция или вид имеет высшую и низшую величину фактора среды, за пределами которых она не может существовать. В двухмерной системе с варьирующими факторами, например, Х и У ниша теоретически будет иметь форму прямоугольника (рис. 1).
Точно так же в трехмерном пространстве ниша будет представлять собой куб (или параллелепипед). При развитии до п варьирующих условий ниша становится гиперобъемом в соответствующей системе координат.
Необходимо, однако, внести два уточнения к этому представлению:
1) Факторы среды взаимодействуют между собой; например, при более высокой температуре повышаются и минимальные требования к влагообеспечению. В таком случае ниша лежит внутри гиперобъема, но не идентична ему.
2) Приспособленность (англ. fitness) популяции не идентична во всех частях ниши. Логично предположить, что она принимает колоколообразную форму при пересечении максимально подходящего пространства ниши или, наоборот, приспособленность разных фенотипов в популяции, сравниваемых в одних условиях, имеет колоколообразное распределение.
Ясно, что организмы любого вида способны нормально расти и развиваться в определенном диапазоне условий внешней среды. При этом степень проявления отдельных факторов или их совокупностей оказывает на растения различное воздействие. В связи с этим различают зоны оптимума, экстремума и промежуточную между ними зону писсимума. Недостаток или избыток какого-либо фактора внешней среды может оказать наибольшее влияние на выживаемость представителей того или иного вида. Границы становятся лимитирующими при приближении к ним какого-либо из условий существования организма. Эта концепция известна как закон минимума Ю. Либиха с 1840 года. Диапазон условий существования организмов, ограниченный лимитирующими факторами (экологическим минимумом и экологическим максимумом), называется пределами толерантности (Ю. Одум, 1986). У каждого вида и сорта растений свои пределы толерантности.
Действие экологического фактора считается стрессовым, если его отклонения в ту или иную сторону от оптимальной величины приводят к гибели 50 % особей в популяции. Зоной оптимума считается такая область условий произрастания, когда нетто-фотосинтез достигает более 90 % своей максимальной величины.
Способность видов занимать разные ниши, характеризующиеся неодинаковой изменчивостью экологических факторов, называют экологической валентностью. Виды растений, адаптированные к узкому диапазону факторов среды, рассматриваются как имеющие низкую экологическую валентность и называются стенотопными. Виды же, способные заселять разные местообитания с изменчивыми условиями, рассматриваются как виды с высокой экологической валентностью и называются эвритопными (Р. Дажо, 1975 — цит. по А. А. Жученко, 1980).
Выделяют три группы экологических факторов: абиотические, биотические и антропогенные. Иногда абиотические и антропогенные факторы объединяют в одну группу.
Абиотические — это факторы, связанные с воздействием на растения косной (или биокосной) природы. К ним относятся климатические (температура, влага, свет, состав воздуха, ветер и т. п.), эдафические, или почвенно-грунтовые (механический состав почвы, ее физические и химические свойства), и орографические (факторы рельефа: высота над уровнем моря, крутизна склона, экспозиция) факторы.
Биотические — это факторы, связанные с жизнедеятельностью биологических организмов (растений, животных, микроорганизмов).
Антропогенные — факторы, связанные с деятельностью человека.
Все эти факторы действуют в комплексе, но с методической точки зрения целесообразно рассмотреть их группы по отдельности.
76.3. Особенности взаимовлияния климатических факторов среды и лесных древесных пород
Тепло
Температура существенно влияет на все физиологические процессы растений (рост, фотосинтез, дыхание, транспирация, транспорт веществ и т. п.). Для каждого вида характерен определенный температурный оптимум существования. На растения отрицательно влияют как очень низкие, так и очень высокие температуры. По отношению к низким температурам различают: холодостойкие растения — переносят длительные низкие температуры: от 0 до 10° С; морозостойкие — не погибают при морозах и переносят промерзание; зимостойкие — переносят как морозы и промерзание, так и другие неблагоприятные условия зимнего периода (иссушение ветром, оттепели и т. п.).
Для условий нашей страны существенное значение имеет отношение растений к морозу и зимним условиям, а также к заморозкам. По отношению к этим факторам разработан целый ряд классификаций, которые приведены в специальной литературе (А. И. Колесников, 1974; И. С. Мелехов, 1980; С. В. Белов, 1983; А. П. Царев, 1997).
Морозостойкость древесных растений связываютсо следующими характеристиками:
а) наличием защитных покровов, ослабляющих промерзание и зимнее иссушение растений (толстая кора, присутствие волосков на побегах и почках, смолистого и воскового налета);
б) способностью переносить в той или иной мере обезвоживание плазмы клеток;
в) интенсивностью накопления защитных веществ (в частности Сахаров);
г) степенью концентрации клеточного сока.
Кроме того, морозоустойчивость связана с сезонным развитием растений. Максимальная устойчивость наблюдается во время органического, или глубокого, покоя. По окончании этой стадии наступает стадия вынужденного покоя, в течение которой даже небольшое потепление провоцирует рост растений, а наступающее после этого похолодание губит их.
Возраст растений также влияет на их морозостойкость. Так, молодые растения более чувствительны к морозам, чем старые.Это, в частности, можно наблюдать на примере ели.
Зимостойкость — понятие более широкое, чем морозостойкость. Оно включает оценку повреждаемости всеми факторами зимы (абсолютные минимумы температур, продолжительность морозов, частоту, амплитуду и продолжительность температурных перепадов, глубину снежного покрова, действие ветра и т. п.). Оценка зимостойкости проводится после перезимовки, в период вегетации. Зимостойкость, как правило, коррелирует с морозостойкостью, однако нередко наблюдаются отклонения, особенно среди интродуцированных растений.
Устойчивость к весенним и осенним заморозкам, по данным Н. Е. Булыгина (1991), тесно связана с фенологическими особенностями растений. От поздневесенних заморозков чаще и сильнее страдают рано вегетирующие виды (ель аянская) или формы (ели европейской, дуба черешчатого, ясеня обыкновенного), виды ранневесеннего цветения, а от ранних осенних — виды, поздно завершающие вегетацию (ольха черная, ива белая), а также поздно распускающаяся форма дуба черешчатого в северной части его ареала (поражаются листья или несозревшие желуди).
Для практических целей оценку отношения древесных пород к низким температурам необходимо проводить в каждом конкретном регионе. На основании таких оценок осуществляется районирование территории для тех или иных пород или сортов.
Вред растениям могут наносить и высокие температуры (опал корневой шейки, ожог коры и т. п.). Способность разных видов растений по-разному реагировать на высокие температуры выражена также довольно отчетливо. Наиболее высокую выносливость к сверхвысоким температурам демонстрируют растения жарких и сухих районов — эуксерофиты, многие суккуленты — кактусы и очитки.
Для жаровыносливых растений характерны некоторые особенности морфологического строения: сильное опушение почечных чешуи и листьев, глянцевитость верхней стороны листьев, их параллельное солнечным лучам расположение и т. п. Растения этой группы обладают высокой жаростойкостью плазмы и другими физиологическими особенностями.
Д. И. Цыганов (1976, 1983) попытался дать сводные шкалы для классификации растений по их отношению к различным экологическим условиям и ввел для этого понятие “экоморфа”. При этом экоморфа определяется как тип отношения растений к экологическим режимам местообитаний. По отношению к температурным условиям экоморфы называются термоморфами (от греч. therme — тепло) или криоморфами (от греч. kryos — холод).
В зависимости от количества поступающего тепла в зоне их обитания, Д. И. Цыганов выделяет следующие термоморфы (ккал/см2 в год):
— гекистотермы — арктическая термозона (0—10);
— субгекистотермы — субарктическая термозона (10—20);
— микротермы — бореальная термозона (20—30);
— субмикротермы — суббореальная термозона (30—40);
— субмезотермы — неморальная термозона (40—50);
— мезотермы — субсредиземноморская термозона (50—60);
— пермезотермы — средиземноморская термозона (60—70);
— субмегатермы — тропическая термозона (70—80);
— мегатермы — экваториальная термозона (80). По отношению к зимним условиям Д. И. Цыганов выделяет следующие криоморфы:
— гиперкриофиты — обитают в поясе очень суровойзимы (средняя температура самого холодного месяца ниже – 32° С);
— перкриофиты — пояс суровой зимы (средняя температура самого холодного месяца от – 24 до – 32° С);
— криофиты — пояс довольно суровой зимы (от – 16 до – 24° С);
— субкриофиты — пояс умеренной зимы (от – 8 до – 16° С);
— гемикриофиты — пояс мягкой зимы (от 0 до – 8°С);
— акриофиты — пояс теплой зимы (от 0 до + 8°С);
– субтермофиты — пояс очень теплой зимы (от + 8 до + 16 °С);
– термофиты — пояс невыраженной зимы (выше + 16 °С). Эти и последующие системы экоморф могут быть использованы при оценке экологических особенностей популяций и форм лесных пород в конкретных географических условиях их произрастания.
Влага
По отношению к влаге (атмосферной и почвенной) выделяют четыре основные группы растений: гидрофиты, гигрофиты, мезофиты и ксерофиты. Кроме того, при большей детализации выделяются оксилофиты и психрофиты (П. Л. Богданов, 1974).
Гидрофиты — это водные семенные растения, полностью или частично погруженные в воду (элодея, кувшинки), а также те, у которых вегетативные органы могут развиваться как в воде, так и в воздухе (стрелолист, лютик водяной). Влияние водной среды очень сильно отразилось на морфологическом и анатомическом строении этих растений. Проводящая система у них развита слабо; кутикула, предохраняющая клетки от высыха ния, отсутствует; механические элементы в тканях не развиты. С другой стороны, у гидрофитов хорошо развиты воздухоносные ткани: воздушные полости у некоторых растений занимают до 70 % объема тела, что необходимо в этой среде обитания; осмотическое давление внутри клеток низкое. Древесных растений среди гидрофитов нет.
Гигрофиты — растения, обитающие во влажных местах. Их корни и корневища находятся в воде или в сильно влажной почве (берега водоемов и рек, сырые луга и сырые леса). Осмотическое давление в клетках этих растений невысокое, корневая система поверхностная, слабо развитая, листовые пластинки большей частью крупные. К гигрофитам относятся некоторые виды мхов, таволга; из древесных растений — ольха черная и многие ивы.
Мезофиты — растения умеренных мест обитания, обеспеченных влагой, но без ее избытка, а иногда и с недостатком. Это наиболее широко распространенная и многочисленная по видовому составу группа растений. Мезофиты при недостатке влаги способны завядать. Это дает им возможность сильно уменьшать испарение и переносить без особого труда непродолжительный недостаток влаги.
При продолжительной сухости воздуха и почвы у некоторых лиственных древесных растений наступает частичный вынужденный листопад:
часть листьев (до 50 %) желтеет и опадает (береза повислая, липа мелколистная, вяз гладкий, акация желтая). При наступлении более влажного периода такие деревья продолжают вегетацию с уменьшенной листовой поверхностью.
Ксерофиты — растения, обитающие в условиях с постоянным или сезонным дефицитом влаги. Для растений необходимо различать физический недостаток воды (физическую сухость), когда в почве воды нет, и физиологический недостаток воды, когда вода в почве есть, но использовать ее растение в достаточной мере не может из-за низкой температуры, сильной засоленности или высокой кислотности. Как физическая, так и физиологическая сухость вызывает у растений в некоторых отношениях одинаковую приспособленность. Однако приспособления против засухи у растений самые разнообразные, так же как разнообразны и типы ксерофитов. Ксерофиты разделяют на две большие группы: эуксерофиты и суккуленты. Наиболее распространена группа эуксерофитов.
Для них характерны:
1) глубокая (до 10 м у саксаула) или интенсивная поверхностная корневая система;
2) уменьшение листовой поверхности или полная редукция листьев (саксаул, джузгун, хвойник, дрок), а также свертывание листьев во время засухи;
3) утолщение наружныхстенок эпидермиса листьев (гультемия);
4) густое опушение листьев (лох, багульник) или восковой налет на листьях;
5) особое строение устьиц, обеспечивающее их плотное закрывание в период засухи;
6) высокое осмотическое давление в клетках (до 80 атм).
Для суккулентов характерно наличие водонакапливающей и водоудерживающей ткани (паренхимы) в стеблях и листьях (кактусы, алоэ и др.).
Оксилофиты — это растения болот с характерной для них высокой плотностью почв, плохой аэрацией, неблагоприятными тепловым и водным режимами, что создает неблагоприятные условия для роста растений. И несмотря на избыток физической влаги, растения ощущают ее физиологический недостаток.
На болотах растут сосна и вечнозеленые кустарники двух типов:
а) брусники: брусника (на микроповышениях), клюква, подбел — растения, имеющие небольшие листья с загнутыми краями, сильно развитой кутикулой и восковым налетом;
б) эрикоидного, или верескового, типа: водяника, вереск (ввиду его экологической пластичности), багульник—растения с мелкими листьями, часто сильно завернутыми и образующими полость, прикрытую волосками.
Психрофиты — растения, произрастающие на холодных почвах (карликовые растения тундр).
По отношению к водному режиму почв Д. Н. Цыганов (1983) предложил следующий ряд гидроморф:
— гиперксерофиты — растения пустынного типа увлажнения почв;
— ксерофиты — растения полупустынного типа увлажнения почв;
— субксерофиты — растения сухостепного типа увлажнения почв;
— семиксерофиты — растения среднестепного типа увлажнения почв;
— субмезофиты — растения лугово-степного типа увлажнения почв;
— мезофиты — растения сухолесолугового типа увлажнения почв;
— пермезофиты — растения влажнолесолугового типа увлажнения почв;
— семигигрофиты — растения сыролесолугового типа увлажнения почв;
— субгигрофиты — растения болотно-лесолугового типа увлажнения
почв;
– гигрофиты — растения болотного типа увлажнения почв;
– субгидрофиты — растения прибрежно-водного типа увлажнения
почв;
– гидрофиты — растения водоемного типа увлажнения почв.
Лесоводами разработан ряд шкал по требовательности древесных пород к влаге, а также по устойчивости к продолжительности затопления
(А. И. Колесников, 1974; И. С. Мелехов, 1980).
Свет
Солнечный свет — практически единственный источник энергии, обеспечивающий рост и существование растительности. Причем свет нужен не только как источник тепла, но и как явление, при наличии которого происходит фотосинтез.
Освещенность на поверхности Земли варьирует в широких пределах. Она зависит от высоты стояния солнца над горизонтом, т. е. от угла падения солнечных лучей, длины дня и условий погоды, от прозрачности атмосферы и других факторов.
Белый солнечный свет представляет смесь различных цветов, определяемых длиной волны 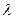 или частотой
или частотой 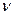 электромагнитных колебаний. Видимые лучи, охватывающие все семь цветов радуги, имеют = 400— 700 нм (м х 10-9) и =7,5 х 1014—4,3 х 1014 Гц. Ближние инфракрасные невидимые лучи имеют = 701—1000 нм и = 4,3х1014—3,0х1014 Гц. Ультрафиолетовые лучи с =200—390 нм сильно поглощаются атмосферой Земли и до ее поверхности доходит только 4—5 % их потока.
электромагнитных колебаний. Видимые лучи, охватывающие все семь цветов радуги, имеют = 400— 700 нм (м х 10-9) и =7,5 х 1014—4,3 х 1014 Гц. Ближние инфракрасные невидимые лучи имеют = 701—1000 нм и = 4,3х1014—3,0х1014 Гц. Ультрафиолетовые лучи с =200—390 нм сильно поглощаются атмосферой Земли и до ее поверхности доходит только 4—5 % их потока.
Видимую солнечную радиацию принято называть фотосинтетически активной радиацией (ФАР). Именно за счет ее протекают процессы фотосинтеза в зеленых растениях. Инфракрасные лучи — это лучи тепловые, зеленая растительность поглощает их мало. При ясном небе доля ФАР (в сумме ФАР + инфракрасная радиация) ниже, при облачном — выше. В среднем соотношение ФАР и инфракрасной радиации принимается равным 50 : 50.
Солнечная радиация делится на отраженную и поглощенную. Так, свежевыпавший снег отражает 90 % радиации, а поглощает 10 %, березовый сомкнутый лес отражает 30 % и поглощает 70 % радиации, а еловый — соответственно 18 и 82 %.
Следует также отметить, что хлорофиллсодержащие растения отражают лучи избирательно. Характерным для них является малое отражение в видимой области спектра ФАР (отражательная способность, или яркость 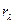 = 3—6 %) и значительно большее — в инфракраснойзоне ( = 44— 70 %).
= 3—6 %) и значительно большее — в инфракраснойзоне ( = 44— 70 %).
Отраженная часть света уходит в пространство, а поглощенная — служит источником для обеспечения жизненных процессов растений: фотосинтеза, транспирации, физического испарения и т. п.
Потребность в свете различных древесных пород неодинакова. Есть древесные породы, которые нуждаются в лучшем освещении и плохо переносят затенение. Такие породы получили название светолюбивых, или гелиофитов (от греч. гелиос — солнце). Некоторые древесные породы могут произрастать в условиях меньшей освещенности, они способны переносить, не погибая, значительное затенение. Их называют теневыносливыми, или сциофитсши (от греч. скиа — тень).
Весь спектр гелиоморфов Д. И. Цыганов (1983) делит на пять групп:
— гелиофиты — тип светового режима открытых пространств;
—субгелиофиты—тип светового режима полуоткрытых пространств (кустарниковых зарослей, опушек, полян, редколесий и т. п.);
— семигелиофиты — тип светового режима светлых лесов (светло-хвойных, мелколиственных, сильно расстроенных темнохвойных и широколиственных);
— субсциофиты — тип светового режима тенистых лесов (темнохвойных и широколиственных средней сомкнутости);
— сциофиты — тип светового режима особо тенистых лесов (некоторых особо высокосомкнутых темнохвойных и широколиственных лесов:
пихтачей, реже ельников, часто бучин, каштанников и т. д.).
Потребность растений в свете, как и отношение к другим экологическим факторам, не является величиной строго постоянной для данного вида, с возрастом она меняется. Так, всходы и молодые деревья обычно более теневыносливы, чем взрослые деревья. Меняется требовательность к свету и с изменением плодородия почвы. На бедных почвах растения становятся более светолюбивыми, чем на почвах более плодородных.
Говоря о значении света для древесных пород, необходимо отметить еще одно явление. Продолжительность солнечного дня постоянна лишь на экваторе. Здесь день, как и ночь, длится 12 часов. Длительность солнечного дня в течение летнего периода возрастает в направлении обоих полюсов; на полюсе целое лето длится полярный день, а зимой — полярная ночь.
Разные группы растений неодинаково реагируют на продолжительность освещения. Одни виды характеризуются как растения длинного дня (молодило, белена, хлебные злаки), другие приспособились к короткому дню (табак, рис, просо, соя, конопля), третьи на длину дня заметно не реагируют (гречиха, горох и др.). Реакция растений на определенную продолжительность дня и ночи называется фотопериодизмом (от греч. фотос — свет и периодос — вращение, круговращение, чередование).
Если чувствительные к фотопериоду растения из районов с коротким днем культивировать в районах с длинным днем, то они развиваются ненормально: не прекращают роста до осенних холодов и страдают или погибают от морозов (акация белая, гледичия трех колючковая в Санкт-Петербурге, сирень на Кольском полуострове и др.). С другой стороны, П. Л. Богданов (1974) отмечает, что у лиственницы при сокращенном дне на три недели раньше прекращалась вегетация, хвоя желтела и опадала.
Шведские исследователи считают, что при выборе экотипов лесных древесных пород для северной Швеции наибольшее значение имеют температурный фактор и фотопериод (A.J.Ola Rosval, 1995; G. Eriksson, I.Ekberg, 1997).
Явление фотопериодизма используется в селекции также для ускорения процессов цветения и плодоношения.
Дата добавления: 2017-09-01; просмотров: 2494;
Поиск по сайту
Узнать еще
- Cтандарт породы Аляскинский маламут FCI
- I.2. Антигены системы АВ0. Генетика. Структура
- III.3. Классификация пород-коллекторов нефти и газа
- XIII. РАЗРАБОТКА ПЛАСТОВ, ОПАСНЫХ ПО ВНЕЗАПНЫМ ВЫБРОСАМ УГЛЯ (ПОРОДЫ) И ГАЗА, И ПЛАСТОВ, СКЛОННЫХ К ГОРНЫМ УДАРАМ
- Імунологія і генетика туберкульозу
- А) Аллопатрическое породообразование
- А) Происхождение и классификация горных пород
- Абердин-ангусская порода

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
