Генетический контроль мейоза
Начиная с 30-х годов 20-го века изучаются вопросы, связанные с контролем мейоза определенными генами. Значение закономерностей генетической регуляции мейоза дает исследователю ключи управления этим сложным и важным процессом в жизни организмов с половым воспроизведением. В век биотехнологии и генной инженерии, давших небывалые возможности для конструирования новых генотипов животных и растений, проблемы мейоза выступают на первый план, поскольку независимо от того, как созданы организмы, их судьба будет зависеть от способности гамет пройти «сито» мейоза и оставить жизнеспособное потомство. В данном случае мейоз выступает в одной из своих эволюционных функций — как барьер на пути нежизнеспособных комбинаций хромосом и генов.
Основные вопросы, решаемые учеными:
Сколько и каких генов вовлечено в мейоз?
Каковы генетические механизмы его запуска?
Как происходит регуляция ключевых событий мейоза?
Каким образом гены контролируют переход от одного события мейоза к другому?
Каков порядок включения генов в ходе мейоза?
Для ответа на поставленные вопросы была разработана методология исследований, основанная на поиске мей-мутантов и создании коллекций.
Такие коллекции были созданы на объектах: Sacchoromyces cerevisiae, Neurospora crassa, Drosophila melanogaster, Pisum sativum, Zea mays. (дрожжи, нейроспора, дрозофила)
Одним из предварительных способов обнаружения мейотических мутантов служит выявление стерильных форм у растений и животных. Затем определяют, не зависит ли стерильность от нарушений в мейотическом делении. Появление анеуплоидных организмов в потомстве указывает на нарушение распределения хромосом в мейозе. По этому показателю открыты гены типа claret у различных видов дрозофилы. Нарушение ожидаемого соотношения отдельных аллелей в потомстве — следующий показатель появления мейотических мутаций.
Следующий этап — это классификация мей-мутантов. Систематизация мей-мутантов показала, что в растительном царстве — от одноклеточной водоросли и грибов до покрытосеменных растений — и в царстве животных — от дрозофилы до человека — существуют однотипные мутации.
1.Благодаря полученным коллекциям мей-мутантов было установлено, что существует строгий генетический контроль семи ключевых цитогенетических событий мейоза, каждый из которых контролируется группой генов, действующих относительно независимо друг от друга. К таким событиям относят:
Готовность клеток к мейозу.
Запуск мейоза.
Регуляция первого деления мейоза (редукция).
Узнавание, сближение и синапсис гомологов.
Образование и поддержание хиазм.
Расхождение гомологичных хромосом.
2. Также установлено, что существует несколько (по крайней мере, три) уровня иерархии генов, управляющих мейозом: гены, контролирующие ключевые блоки мейоза; элементарные события в пределах блока; особенности поведения хромосом.
3.Ступенчатое включение мей-мутаций в процессе мейоза.
Эволюция мейоза
Эволюция мейоза в жизни организмов неразрывно связана с возникновением полового процесса. Появление различных типов мейоза указывает на неоднозначное становление этого сложного и столь важного процесса в жизни высших организмов.
Мейоз с одним делением ядра — под этим понятием понимается спаривание, а затем расхождение гомологов без предварительной их редупликации. Биваленты при таком мейозе не четыреххроматидные, а двуххроматидные, хиазмы не возникают. Весь процесс хромосомной редукции завершается в одном делении. Если за ним и будет следовать второе деление, оно окажется чисто митотическим с обычной предшествующей ему редупликацией хромосом.
Одноступенчатый мейоз впервые описан Э. Кливлендом у жгутиконосцев — кишечных паразитов таракана. Наиболее существенная черта одноступенчатого мейоза — это отсутствие удвоения хромосом в мейотической профазе. Именно это делает невозможным образование хиазм и кроссинговер. При одноступенчатом мейозе не происходит предмейотической репликации ДНК.
Исключение 1. У некоторых низших Protozoa имеет место одноступенчатый мейоз, при котором кинетохор в начале анафазы не расщеплен, но в ходе анафазы он расщепляется, и тогда идет каскадный процесс разъединения сначала гомологов, а затем сестринских хроматид каждого гомолога.
Исключение 2. Это мейоз без синаптонемного комплекса, встречается у грибов S.pombe и A.nidulans. В профазе I мейоза у S.pombe формируются линейные элементы, которые затем не превращаются в латеральные элементы СК.
Исключение 3. Это мейоз у самцов дрозофиллы, при котором редукция числа хромосом не происходит без СК, кроссинговера и хиазм. Вместо хиазм гомологичные хромосомы в метафазе I мейоза у самцов на короткое время соединяются белковыми волокнистыми структурами — коллахорами. У самцов и самок одного вида функционируют разные механизмы мейоза.
2.Мейоз с двумя делениями ядра. В мейотической профазе происходит конъюгация хромосом. В диакинезе и в метафазе первого мейотического деления биваленты сильно укорачиваются, в анафазе — разъединяются на диады. В интеркинезе диады имеют характерную четырехплечную форму; в метафазе второго деления хроматиды диад разъединяются. Таким образом, мейоз полностью укладывается в классическую схему.
У простейших можно наблюдать образование синаптонемного комплекса, где просматриваются боковые элементы и центральный элемент.
Эволюционные преимущества двухступенчатого мейоза очевидны: он обеспечивает значительно большую частоту наследственной рекомбинации по сравнению с одноступенчатым: если в одноступенчатом мейозе происходит рекомбинация только тех генов, которые относятся к разным группам сцепления (за счет независимого расхождения хромосом в анафазе), то в двухступенчатом мейозе происходит рекомбинация генов, относящихся и к одной, и к разным группам сцепления (т.к. осуществляется кроссинговер).
Мейоз у отдаленных гибридов
Основу цитогенетических исследований отдаленных гибридов составляет изучение конъюгации хромосом в процессе мейоза.
В процессе отдаленной гибридизации у гибридов можно встретиться с двумя ситуациями.
Первая: родительские формы, несмотря на различие в генах, имеют «соответствующие» хромосомы, которые могут конъюгировать, не снижая жизнеспособности и фертильности гибридов. К этой группе относятся скрещивания некоторых рас, разновидностей и видов. Такие скрещивания называются конгруэнтными.
Вторая: родительские формы имеют «несоответствующие» хромосомы или их разное число, в результате чего у гибридов возникают нарушения в мейозе.
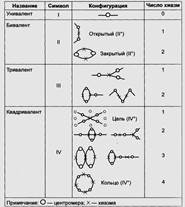
В зависимости от характера конъюгации хромосом выделяют несколько типов конфигурации хромосом в мейозе:
Биваленты — конфигурации, состоящие из двух хромосом. Открытые биваленты соединены одной хиазмой, закрытые — двумя.
Триваленты — конфигурация из трех хромосом, соединенных двумя хиазмами.
Квадривалент — конфигурация, состоящая из четырех хромосом. Выделяют несколько типов квадривалентов, однако наиболее часто встречаются квадриваленты открытые и закрытые.
Хромосомы, которые не нашли партнера, являются одиночными и называются униваленты (табл. 2).
Таблица 2. Типы конфигураций хромосом в мейозе
В процесс конъюгации у отдаленных гибридов могут вступать несколько типов хромосом:
Гомологичные — полностью одинаковые хромосомы.
Гомеологичные хромосомы — это хромосомы, у которых последовательность локусов чаще всего нарушена, вследствие чего их конъюгация затруднена или отсутствует.
Типы нарушений в мейозе у отдаленных гибридов (рис. 9):
1.Неравномерное расхождение хромосом к полюсам в анафазе I.
Образование женских и мужских гамет с разным числом хромосом и, как следствие, различная жизнеспособность гамет. Чаще гибнут мужские гаметы с несбалансированным числом хромосом, а при нормальном оплодотворении появляются анеуплоидные растения.
2.В анафазе I задержка унивалентов (одного или нескольких) на экваторе.
3.Образование микроядер в диаде микроспор и макроспор, образование хромосомных и хроматидных мостов, а также нарушение функций веретена деления.
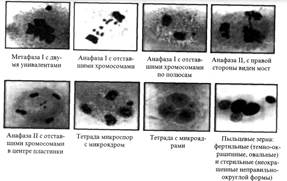
Рис. 9. Типы нарушений в мейозе у отдаленных гибридов
4.Нарушение процесса мейоза во время второго деления.
Появление отставания хромосом, мостов, фрагментов, многополюсность.
5.Часто в пределах одного пыльника встречаются клетки, находящиеся в стадиях от диакинеза до тетрад, наблюдается асинхронность деления.
6.Возникновение дефективных тетрад. Это тетрады с различным числом микроядер, образование полиад - пентад, гексад.
Дата добавления: 2017-03-12; просмотров: 4700;
Поиск по сайту
Узнать еще
- II. Качественный контроль (социологический анализ).
- XXIV. КОНТРОЛЬ РУДНИЧНОЙ АТМОСФЕРЫ
- Автоматическая защита и контроль работы холодильных установок.
- Автоматические контрольные устройства
- Автоматический контроль формы деталей.
- Активный контроль заготовок до обработки. Блокирующие устройства
- Альтернатива турникетам – перронный контроль.
- Анализ и контроль маркетинга

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
