Что такое Матричный биосинтез
Это одна из интересных проблем молекулярной биологии, где много еще таких механизмов нерасшифрованно. В живом организме постоянно происходит наряду с распадом синтез белка. Метод линейных атомов позволил установить, что в состав клеток входит большое количество разнообразных белков и скорость синтеза их различны. Белки эритроцитов обмениваются в течении 2-3 месяцев, в тоже время белки уже обмениваются очень быстро, установлено, что основные белки нервной ткани обмениваются в течение 21 дня.
Белки в клетках органов и тканей вступает во взаимодействие с различными компонентами и поэтому в клетках должен существовать механизм, который бы обеспечивал безошибочный синтез белковых веществ. Это имеет значение для метаболических процессов.
Среди заболеваний связанных с нарушениями синтеза белка можно называть «альбинизм». Что происходит:
1) Нарушение процесса образования пигмента меланина, он вырабатывается в специальных клетках меланоцитах, которые находятся в коже, в волосных луковицах, сетчатке глаза. Прекращается выработка пигмента вследствие нарушения процесса превращения фенилаланина в тирозин. При альбинизме не вырабатывается фермент – тирозиназа. Он способствует образованию в дальнейшем пигмента меланина.
Признаки: молочно белый цвет кожи, светлые волосы, светлая радужной оболочки, депигментизация сетчатки, снижение остроты зрения (люди страдают, но живут)
2) Серповидноклеточная анемия происходит вследствие замены одной аминокислоты глу на вал и гемоглобин принимает форму серпа и не может выполнять свою функцию основную – транспорт О2
Для того чтобы процесс биосинтеза белка проходил нормально необходимо:
1) Поток материи (аминокислоты из которых будут строится белки), обязательное присутствие незаменимых аминокислот. Поток должен быть как количественным так и качественным. Если с пищей наступает недостаточное количество незаменимых аминокислот, то наблюдается белковое голодание. Это приводит к нарушению азотистого равновесия (он становится отрицательный). Это важно учитывать при составлении рационов питания;
2) Поток энергии. Установлено, что синтез сложных веществ в организме протекают с потреблением источников энергии – энергии АТФ, ГТФ и и.д.;
3) Необходима информация о том, какой белок должен синтезироваться;
4) Необходимы непосредственные участники синтеза белка – различные типы РНК, позволяющие клетке синтезировать заданный белок. РНК – переносчик потока информации от ДНК к месту синтеза белка.
Начнем с общих механизмов синтеза ДНК
1) Корнберг в 1953 году предложил энзиматическим путем в безклеточной среде с участием ДНК - полимеразы
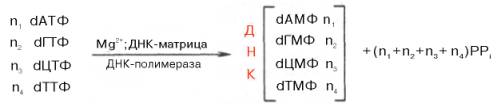
Открытие в 1960 одновременно в 2х лабораториях США фермента РНК полимеразы, катализирующего синтез РНК из свободных нуклеотилов. Способствовало расшифрованию механизма синтеза РНК.
Наиболее изучена РНК – полимераза прокариот Е.coli с АС 487000 состоит из 5 субъединиц.
РНК – полимеразы (называются ДНК – зависимой полимеразой) было установлено, что молекуле ДНК необходимы не только для реакции полимеризации, но что она определяет последовательность рибонуклеотидов во вновь синтезируемой молекуле РНК с заменой тимизинового нуклеотида ДНК на уридиловой в РНК. В общем еще синтез РНК можно представить так:
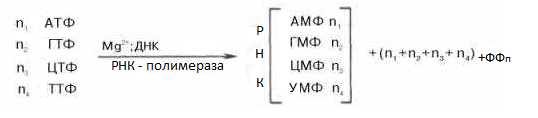
У Е. coli предполагают, имеется единственная ДНК зависимая РНК – полимераза, которая синтезирует все типы клеточных РНК. Менее изучены РНК – полимеразы эукариотов. Из клеток животных выделены 3группы РНК – полимераз А, В, С, которые принимают участие в синтезе соответственно рРНК, мРНК и тРНК.
Матричный биосинтез состоит из 3х этапов:
1. Биосинтез ДНК – репликация (механизм удвоения ДНК), репарация (ферментативные механизмы, обнаруживают и исправляют повреждения ДНК)
2. Транскрипция – биосинтез ДНК (тРНК, рРНК, мРНК)
3. Этап биосинтеза белка – трансляция
Биохимический смысл процессов репликации заключается в том, что они протекают в несколько этапов. (рис.1)
На первом этапе - инициации - происходит образование с участием ферментов (ДНК -хеликаз, ДНК - гираз) репликационных вилок, т.е. если мы имеем 2-х цепочную ДНК, то на определенном этапе одна из цепочек откручивается и ушедшая часть достраивается в виде антипараллельной цепи (рис. 1 ).
При инициации к цепям ДНК последовательно присоединяются ДНК - связывающие и ДНК - раскручивающие белки, а затем комплексы ДНК - полимераз и ДНК-зависимая РНК – полимераза (праймаза).
Второй этап. Процесс репликации ДНК подвергаются одновременно обе цепи. Рост дочерних цепей осуществляется в направлении
5’ _____3’. Первая стадия осуществляется при помощи ДНК - полимеразы 111
далее принимает участие ДНК - полимераза 11 .Синтез на одной цепи идет не прерывно, а на другой фрагментарно (фрагменты Оказаки). Вторая стадия завершается отделением праймеров, объединением отдельных фрагментов ДНК при помощи ДНК - лигаз и формированием дочерней цепи ДНК.
Третий этап - терминация синтеза ДНК, наступает в результате обрыва цепи за счет исчерпывания ДНК матрицы. Точность репликации велика. Если будет ошибка, то она может быть исправлена в ходе репарационных процессов.
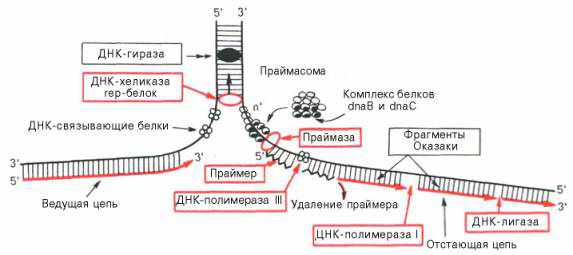
Рис.1 Схема основных этапов репликации ДНК (по Т.Т.Березову и Б.Ф.Коровкину)
Репарация ДНК и РНК.
Ряд экзогенных и эндогенных факторов приводят к различным повреж –дениям ДНК в клетке. В клетке существуют системы репарации ДНК. Это ферментативные механизмы, которые обнаруживают и исправляют повреждения.
Какие необходимы для этого условия?
1.Необходимо узнавание места повреждения ДНК (с помощью эндонуклеаз);
2.Удаление поврежденного участка (с помощью ДНК –гликозидаз);
3.Синтез нового фрагмента (ДНК – полимеразе репарирующая);
4.Соединение образования новых участков со старой цепью (фермент ДК -лигаза).
Транскрипция РНК.
Транскрипция отличается от репликации. При репликации реплицируется полностью одна из цепей ДНК, а при транскрипции транскрибируется
отдельные гены. Поэтому каждый ген ДНК несет свою информацию.
Процесс образования мРНК на ДНК - затравке возможен только на функционирующей ДК. Процесс транскрипции - многоступенчатый. До открытия феномена сплайсинга (созревание, сращивание) мРНК было известно, что многие мРНК эукариот синтезируются в еще гигантских высокомолекулярных предшественников (пре - мРНК), которые уже в ядре подвергаются посттранскрипционному процеосингу. Оказалось, что ген у эукариотов имеет сложное мозаичное строение. Он включает в себя участки, несущие информацию, это кодирующие - экзоны и участки не несущие информации, т.е. ничего не кодирующие - интроны . Отсюда и возникло понятие об экзонинтронной структуре (рис. 2).
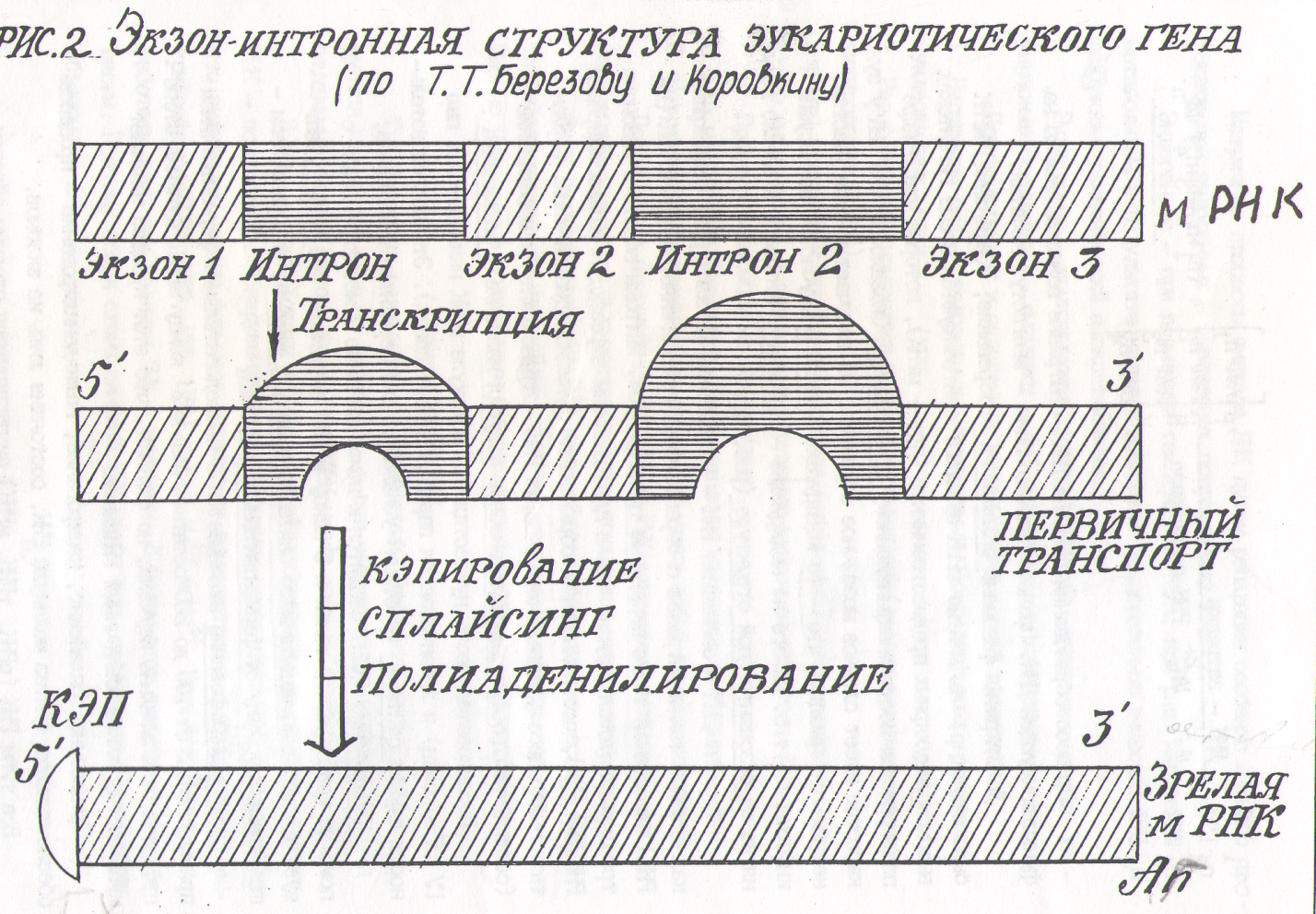
Фермент ДНК - зависимая РНК - полимераза катализирует транскрипцию как экзонов так и интронов с образованием гетерогенной ядерной РНК (гя РНК) называемой также первичным транскриптом. Интроны вместе с экзонами транскрибируются; однако еще в ядре интроны вырезаются малыми ядерными РНК (мя РНК), что приводит к образованию функционирующей мРНК. Ферментативный процесс удаления интронов из РНК - транскрипта и объединение (соединение), соответствующих экзонов получил название - сплайсинга .
Последовательность нуклеотидов в молекуле мРНК начинается с пар ГУ (5'- конец) и заканчивается парой АГ (3' - конец). Эти последовательности служат сайтами (местами) узнавания для ферментов сплайсинга.
Кэпирование (КЭП) сводится к присоединению 7 метилгуанозина с помощью трифосфатной связи к 5' концу мРНК, считают, что "НЭП" участвует в узнавании подходящего сайта на молекуле мРНК и, возможно, защищает саму молекулу от ферментативного распада.
Полиаденилирование заключается в последовательном ферментативном присоединении от 100 до 200 остатков АМФ к 3' концу мРНК. Функция этого процесса окончательно изучена, но считают, что этот процесс защищает мРНК от гидролиза клеточными РНКазами.
Процессинг, сплайсинг, кэпирование, полиаденилирование - процессы обеспечивающие синтез молекулы РНК, состоящие лишь из экзонов.
Все типы РНК (рРНК, тРНК, мРНК) синтезируются сходным образом.
Поэтому для любой молекулы РНК, имеющейся в организме можно найти участок ДНК, которому она комплементарна. Но все же в синтезе различных видов имеются некоторые особенности.
мРНК - синтезируется гораздо большего размера, чем требуется для синтеза белка. Так белок иммуноглобулин включает тяжелую цепь, кодируется 1800851 нуклеотидными остатками, из них непосредственно структуру белка кодирует 1300 нуклеотидных остатка.
тРНК - синтезируется также как и мРНК, но при этом синтез идет из большего предшественника. Этот процесс подвергается сплайсингу при участии ферментов цитоплазмы.
рРНК - бывает нескольких типов. У прокариотов синтез рРНК трех типов 235, 16S , 5S . Они образуются из длинного предшественника пре - рРНК. Из них идет образование одной из субъединиц рибосомы.
Таким образом транскрипция - многоступенчатый процесс, в результате которого синтезируются все виды РНК.
Биосинтез белка (трансляция).
Генетический текст при трансляции переводится в линейную последовательность аминокислот полипептидной цепи белка.
Процесс трансляции можно разделить на два этапа, которые имеют разную локализацию в клетке: рекогниция (узнавание аминокислот) и собственно биосинтез белка. Рекогниция протекает в цитоплазме, а биосинтез белка протекает в рибосомах.
Рекогниция,или узнавание аминокислот. Сущность узнавания аминокислот состоит в том, чтобы соединить аминокислоту со своей тРНК. Структура тРНК обладает качествами потенциального "переводчика", так как в одной молекуле совмещены способности ''читать'' нуклеотидный текст (антикодон тРНК специфически спаривается с кодоном мРНК и нести (на акцепторном конце) свою аминокислоту. Специальные ферменты обеспечивают узнавание тРНК своей аминокислоты. Эти ферменты получили название аминоацил - тРНК - синтетаз (АРСазы). Аминокислоты при этом должны быть активированы, активация осуществляется также при помощи АРСаз. Этот процесс протекает в 2 стадии:
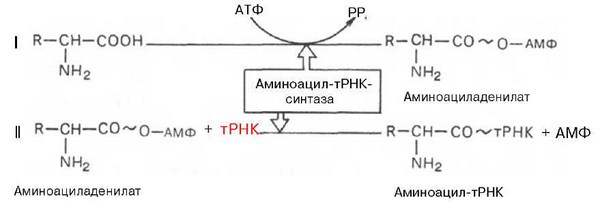
Рибосомы, не участвующие в синтезе белка легко диссоциируют на субъединицы. В клетке рибосомы или находятся в свободном состоянии или связаны с мембранами эндоплазматической сети. Свободное перемещение рибосом в различные участки клетки или соединение их в разных местах с мембранами эндоплазматического ретикулума, очевидно, дает возможность собирать белки в клетке там, где это нужно.
Биосинтез белков отличается от других типов матричных биосинтезов-репликации и транскрипции - двумя особенностями:
1) Нет соответствия между числом знаков (мономеров) в матрице и продуктов реакции в мРНК 4 разных нуклеотида, в белке 20 разных аминокислот;
2) Структура рибонуклеотидов (мономеров матрицы) и аминокислот (мономеров продукта) такова, что между мРНК (матрицей) и полипептидной цепью белка (продуктом) нет комплементарности.
Синтез белка или трансляцию делят на 3 фазы: инициацию (начало), элонгацию (удлинение полипептидной цепи), терминация (окончание).
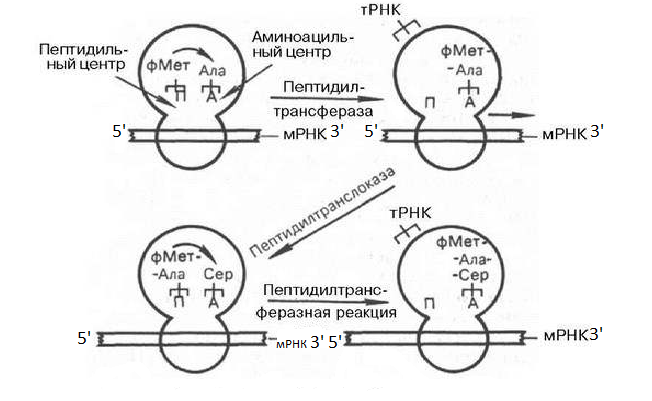
В настоящее время установлено, что для начала синтеза белка существует специальный инициирующий комплекс (формил мет тРНК и мРНК связанные с несколькими молекулами белка ГТФ). Происходит взаимодействие между кодонами мРНК и антикодонами формил мет РНК. (рис.3)
Вначале инициирующая формил мет РНК связывается с большой субъединицей рибосомы в участке П (пептидильный центр). Следующая аминокислота в виде алат РНК связывается в участке А (аминоацильный центр). Рибосомы за счет взаимодействия антикодона ала тРНК и кодона мРНК. В результате «NH2» этой аминокислоты оказывается вблизи от «СООН» группы первой аминокислоты с помощью пептидотрансферазы образуется пептидная связь в участке А. Образовавшийся дипептид переносится транслоказой из участка А в участок П, вытесняя оттуда тРНК, которая вновь может вступать во взаимодействия с другой аминокислотой, необходимо участие ГТФ. Под действием пептидтрансферазы пептидная цепь с учатска П переносится на участок А. Рибосома сдвигается и против А участка становится новый кодон мРНК. На этом один рибосомальный цикл завершается. Процесс синтеза белка продолжается до тех пор, пока к А участку не подойдет бессмысленный кодон (УАГ, УАА, УГА). На этом синтез белка заканчивается и синтезируемый пептид с участка П отделяется от поверхности рибосомы.
Большинство синтезируемых белков остается в клетке, а часть уходит путем экзоцитоза. Для этого требуется энергия АТФ, поэтому при дефиците АТФ белки задерживаются в клетке. Особенно активно белки выделяются железистыми клетками и клетками печени. Что происходит дальше с синтезируемым белком?
После отделения от рибосомы она тут же гидролизуется цитоплазматическими рибонуклеазами. Уже в ходе трансляции белок начинает укладываться в трехмерную структуру, которую он окончательно принимаем после отделения синтезированного белка от рибосомы. В результате трансляции не всегда образуется функционально активный белок. Во многих случаях необходимы дополнительные посттрансляцивнные изменения. Например, инсулин, образуется из предшественников (проинсулина) в результате отщепления части пептидной цепи под действием специфических протеаз. Сходным образом, т.е. путем частичного протеолиза, активируются многие проферменты.
Присоединение простетической группы с образованием сложных белков и объединение протомеров олигомерных белков также относятся к посттрансляционньм изменениям. В некоторых белках после завершения синтеза пептидной цепи происходит модификация аминокислотных остатков, например превращение пролина и лизина в гидроксилизин и гидроксипролин в коллагенах, метелирование аргинина и лизина в гистонах, иодирование тирозина в трио глобулине. Некоторые белки подвергаются гликозилированию, присоеди -няя олигосахаридные остатки (образование гликопротеинов). Одной из постсинтетических модификаций является фосфорилирование некоторых остатков тирозина в молекуле белка и в настоящее время рассматривается как один из специфических этапов формирования онкобелков при малигнизации нормальных клеток. Хотя биосинтез белка, представляющий сложный многоступенчатый процесс, однако структурно - функциональные взаимоотношения различных его этапов еще недостаточно изучены.
Рис.3 Схема элонгации полипептидной цепи
| <== предыдущая лекция | | | следующая лекция ==> |
| Энзимодиагностика и энзимотерапия. | | | Переваривание углеводов |
Дата добавления: 2017-03-12; просмотров: 3179;
Поиск по сайту
Узнать еще
- B Убедитесь что звезды распредвалов должным образом точно выставлены до установки и затяжки болтов
- I. Что такое смерть? И что такое жизнь, концом которой является смерть?
- III. Биосинтез крахмала и гликогена.
- Iii. строение, биосинтез и биологическое действие гормонов
- Активация аминокислот при биосинтезе белка.
- Анаболизм прокариот. Биосинтез углеводов
- Билет №8 В. 1. Что такое локализация пожара.
- Биосинтез аминокислот

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
