Типы взаимодействия неаллельных генов: комплементарность, эпистаз, полимерия. Гены – модификаторы.
Фенотип организма формируется под влиянием большого количества генов, а также в результате их взаимодействия.
Все многообразие межгенных взаимодействий можно разделить на две группы: взаимодействие аллельных и неаллельных генов.
1. Аллельные гены находятся в идентичных локусах гомологичных хромосом, и взаимодействие между ними проявляется в форме полного, неполного доминирования и кодоминирования.
2. Неаллельные гены локализованы в разных парах гомологичных хромосом или в одной паре гомологичных хромосом, но в разных ее локусах.
Выделяют три основных типа взаимодействия неаллельных генов.
1. Комплементарность – тип неаллельного взаимодействия генов, при котором сочетание в генотипе доминантных аллелей обоих генов обуславливает появление нового признака.
Впервые подобный тип взаимодействия был изучен У. Бетсоном и Р. Пеннетом у душистого горошка.
При скрещивании двух линий с белыми цветками в F1 дигетерозиготные растения АаВв имели пурпурные цветки, а в F2 было получено 9/16 (A-B-) растений с пурпурными цветками, и 7/16 (3/16 A–bb+ 3/16 aaB– + 1/16 aabb) с белыми, т.е. расщепление составило:
· 9:7.
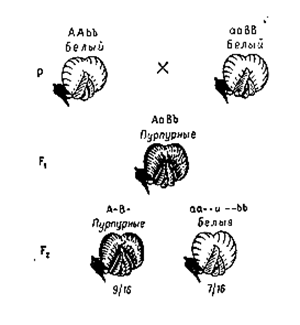
Наследование окраски цветков у Lathyrus odoratus при взаимодействии двух пар генов
Таким образом, взаимодействие доминантных генов А+В обусловливает пурпурную окраску цветков.
При комплементарном взаимодействии генов возможны отклонения от стандартной формулы расщепления по фенотипу (9:3:3:1) при дигибридном скрещивании, а именно:
· 9:6:1
Вариант такого взаимодействия генов характерен для наследования формы плодов у тыквы.
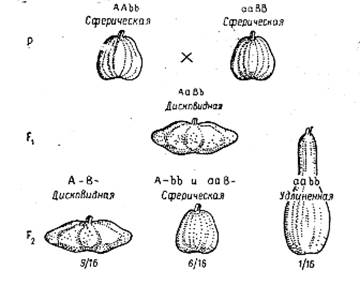
Наследование формы плода у Cucurbita pepo при взаимодействии двух пар генов
У тыквы наблюдается три разновидности плодов: дисковидная, сферическая и удлиненная, причем сферическая форма является рецессивной по отношению к дисковидной.
При скрещивании двух сортов тыквы со сферическими плодами получаются растения F1 с дисковидной формой плодов. В потомстве этих растений в F2 появляются три фенотипических класса в соотношении 9/16 с дисковидными плодами (А–В–), 6/16 – со сферическими (3/16 A–bb+3/16 aaB–) и 1/16 с удлиненными (aabb). Это свидетельствует о том, что каждый из доминантных неаллельных генов А и В детерминирует сходный фенотип – сферическую форму плодов, взаимодействие их доминантных аллелей в генотипе обусловливает дисковидную форму плодов, а взаимодействие рецессивных аллелей - удлиненную форму.
· 9:3:3:1
Подобное расщепление по фенотипу в F2 наблюдается при наследовании окраски глаз у дрозофилы.
При скрещивании линий дрозофилы с ярко-красными и коричневыми глазами получены гибриды F1 с красными глазами.
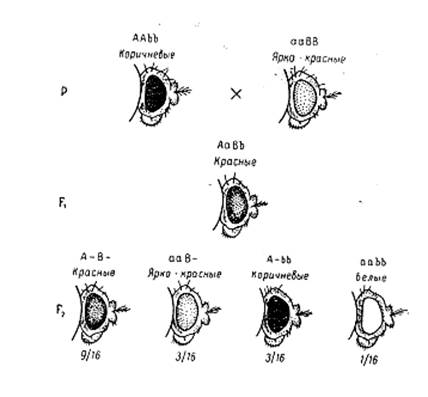
Наследование окраски глаз у Drosophila при взаимодействии двух пар генов
В F2 присутствие доминантных генов А и В у 9/16 особей приводит к формированию красной окраски глаз. Присутствие гена А в гомо- или гетерозиготном состоянии при рецессивном b дает ярко-красную окраску у 3/16 особей, а гены aaB– у 3/16 потомства дают коричневую окраску. Гомозиготы по обоим рецессивным генам aabb (1/16) имеют новый фенотип – белую окраску глаз.
Итак, взаимодействие доминантных генов в генотипе изменяет окраску глаз. Каждый из комплементарных доминантных генов имеет собственное фенотипическое проявление, а двойная рецессивная гомозигота отличается от них по фенотипу.
· 9:3:4
Вариант подобного взаимодействия комплементарных генов можно рассмотреть на примере наследования окраски луковицы. У лука скрещивание формы, имеющей неокрашенную (белую) луковицу, с формой, имеющей желтую луковицу, дает в F1 растения с красной луковицей. А в F2 появляются растения с красной (9/16), желтой (13/16) и белой (4/16) луковицами:
P: ccRR × CCrr
Белая Желтая
Гаметы: cR Cr
F1 CcRr
Гаметы: CR, Cr, cR, cr
F2 9/16 C–R– : 3/16 C–rr : 4/16 (3/16 ccR– + 1/16 ccrr)
Красная Желтая Белая
Красная окраска луковицы обусловлена наличием двух доминантных генов (С–R–). Доминантный аллель Сдетерминирует желтую окраску луковицы, а рецессивный аллель с– белую. Доминантный ген R не имеет собственного фенотипического проявления и объединяется по фенотипу с рецессивной гомозиготой гена с, аллель r не влияет на проявление окраски.
Таким образом, комплементарными являются гены, которые при совместном действии в генотипе в гомо- и гетерозиготном состоянии (А–В–) обусловливают развитие нового признака. Действие каждого гена в отдельности (А–вв или ааВ–) воспроизводит признак лишь одного из скрещиваемых родителей.
Расщепление в F2 по фенотипу может быть разнообразным: 9:7, 9:6:1, 9:3:3:1, 9:3:4.
2. Эпистаз – тип неаллельного взаимодействия генов, при котором ген одной аллельной пары подавляет действие генов другой пары.
Гены, подавляющие проявление других генов, называются супрессорами, а подавляемые гены – гипостатичными. Выделяют два типа эпистаза: доминантный и рецессивный. При доминантном эпистазе – супрессии ингибирующее действие оказывает доминантный аллель: А>B.
· 13 : 3
Окраска оперения кур определяется двумя генами, взаимодействующими по типу доминантного эпистаза.
Ген С обусловливает окрашенное оперение, ген I подавляет проявление пигмента(I>C); ген с детерминирует белое оперение, ген i на окраску не влияет.
При скрещивании куриц породы леггорн (ССII) с петухами породы белый виандот (ссii) в F2 13/16 кур с белым оперением и 3/16 с окрашенным оперением, у которых нормальный синтез пигмента и проявление гена С не ингибируется эпистатичным геном I.

Дата добавления: 2017-03-12; просмотров: 3848;
Поиск по сайту
Узнать еще
- I. Политический режим: понятие, сущность и основные типы.
- I. Понятие и типы политических партий.
- I.2. Антигены системы АВ0. Генетика. Структура
- II группа действий. ВЗАИМОДЕЙСТВИЯ
- II. Типы политических лидеров.
- II.1.2. Гены. Структура антигенов
- VI. ЩЕЛЕВЫЕ И СПЕЦИАЛЬНЫЕ ТИПЫ АНТЕНН
- А) Типы и конструктивные особенности баз.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
