Общая характеристика оплодотворения
После своего выхода из гонад как яйцеклетка, так и спермий обречены на гибель в течение считанных часов, если они не отыщут друг друга и не сольются в единое целое в ходе процесса оплодотворения. Впоследствии, активированная проникновением сперматозоида, яйцеклетка приступает к осуществлению своей программы развития.
Оплодотворение — слияние сперматозоида с яйцеклеткой, завершающееся объединением их ядер в единое ядро оплодотворенного яйца (зиготы). У подавляющего большинства животных при нормальном развитии именно оплодотворение служит толчком к выходу яйцеклетки из анабиотического состояния, в котором она находится на последнем этапе стадии созревания.
Готовность яйцеклеток к оплодотворению определяется выделением направительных телец. Сперматозоиды проникают в яйцеклетку на разных этапах этого процесса: до выделения (круглые черви), в момент выделения (млекопитающие, включая человека) и после их выделения (иглокожие).
Оплодотворение осуществляет две разные функции:
• половую (комбинирование генов двух родителей) — включает передачу генов от родителей потомкам;
• репродуктивную (создание нового организма) — включает инициацию в цитоплазме яйца тех реакций, которые позволяют продолжать развитие.
Важная роль в процессе оплодотворения принадлежит сперматозоиду, он необходим:
• для активации яйца, побуждения его к началу развития (данная функция неспецифична: в качестве активирующего фактора сперматозоид может быть заменен рядом физических или механических агентов, см. также «Партеногенез»);
• внесения в яйцеклетку генетического материала отца.
Существует несколько принципов классификации процесса оплодотворения:
по месту проникновения сперматозоида в яйцеклетку:
• наружное (оплодотворение происходит во внешней среде);
• внутреннее (оплодотворение происходит в половых путях самки);
по количеству сперматозоидов, участвующих в оплодотворении:
• моноспермное (один сперматозоид);
• полиспермное (два и более сперматозоидов).
У ряда беспозвоночных, рыб, хвостатых амфибий и птиц возможна полиспермия, когда в яйцо проникает несколько сперматозоидов, однако с ядром яйцеклетки сливается ядро одного спермия.
Продолжительность жизни сперматозоидов и яйцеклеток относительно невелика как при наружном, так и при внутреннем оплодотворении. Яйцеклетки многих беспозвоночных, а также рыб и амфибий должны быть оплодотворены сразу же после овуляции.
К моменту встречи с яйцеклеткой сперматозоид должен сохранять не только активное движение, но и свою оплодотворяющую способность. Как правило, они теряют ее раньше, чем способность к движению. Так, у морского ежа оплодотворяющая способность сперматозоидов сохраняется в течение 30 часов, а жизнеспособность в течение двух суток.
Оплодотворяющая способность спермы зависит от многих факторов: концентрации в ней сперматозоидов, температуры, pH среды (в щелочной среде спермии более активны, но быстро растрачивают энергию и раньше погибают, а в кислой их активность меньше, но продолжительность жизни больше).
Конкретные особенности оплодотворения очень сильно варьируют у различных видов. Взаимодействие половых клеток (гамет) канонически подразделяют на четыре стадии (фазы):
• дистантные взаимодействия;
• контактные взаимодействия;
• проникновение сперматозоида в яйцеклетку;
• слияние генетического материала.
Дистантные взаимодействия гамет
Осуществляются на некотором расстоянии, до соприкосновения гамет друг с другом. Они направлены на повышение вероятности встречи сперматозоидов и яйцеклетки. Характерны для водных организмов с наружным типом оплодотворения. При этом животные сталкиваются со следующими проблемами:
• осуществление встречи спермиев и яиц при их низкой концентрации в среде;
• предотвращение оплодотворения яиц спермиями другого вида.
В ходе эволюции выработалось соответственно два механизма для решения поставленных задач. Это видоспецифичное привлечение спермиев и видоспецифичная их активация.
Уже в ходе ранних исследований процесса оплодотворения возникло представление о гамо`нах — веществах, обеспечивающих активацию или блокирование отдельных его этапов.
Гамоны
(от греч. gámos — брак) вещества, выделяемые половыми клетками и способствующие оплодотворению ( Оказывая специфическое действие на Гаметы своего и противоположного пола, Г. контролируют их встречу и содействуют соединению сперматозоида с яйцом. Впервые Г. обнаружены у морского ежа в 1911 Ф. Лилли. Термин «Г.» предложен в 1940 нем. учёными М. Хартманом и Р. Куном. Вещества, выделяемые женскими и мужскими гаметами, названы ими соответственно гиногамонами и андрогамонами. Г. найдены у некоторых растений (водоросли, грибы) и многих животных (моллюски, кольчатые черви, иглокожие, хордовые).
Яйцеклетка способна продуцировать гиногамоны, а сперматозоид — андрогамоны. Полагали, что гиногамон I — низкомолекулярное вещество небелковой природы, которое активирует движение сперматозоидов (повышая тем самым вероятность их встречи с яйцом), преодолевая действие андрогамона I, подавляющего подвижность спермия. Гиногамон II, часто именуемый фертилизином, — гликопротеин, расположенный в периферической области яйца и вызывающий связывание его со сперматозоидами при взаимодействии с комплементарным ему андрогамоном II (антифертилизином), встроенным в поверхностную оболочку спермия, способным разжижать студенистое вещество и растворять оболочку яйца, из-за чего его зачастую отождествляют с гиалуронидазой. Гипотезы оплодотворения, основанные на идее о взаимодействии фертилизина с комплементарным ему антифертилизином, сыграли положительную роль, поскольку позволили обнаружить существование целого семейства специфических молекул на поверхности взаимодействующих гамет (см. далее).
Видоспецифичное привлечение спермиев доказано для многих животных: кишечнополостных, моллюсков, иглокожих и первичнохордовых. Оно представляет собой род хемотаксиса — движения по градиенту концентрации какого-либо вещества. В 80-х гг. XX века удалось идентифицировать два видоспецифичных аттрактанта сперматозоидов морских ежей — сперакт и резакт. Оба вещества, выделенные из студенистой оболочки яиц, относятся к пептидам и содержат 10 и 14 аминокислотных остатков соответственно. При взаимодействии со спермиями они стимулируют их метаболизм и увеличивают подвижность мужских гамет.
Контактные взаимодействия гамет
Как следует из названия, данная стадия начинается с момента контакта сперматозоида с третичной оболочкой яйцеклетки.
9 Первым ее этапом является акросомная реакция (рис. 12).
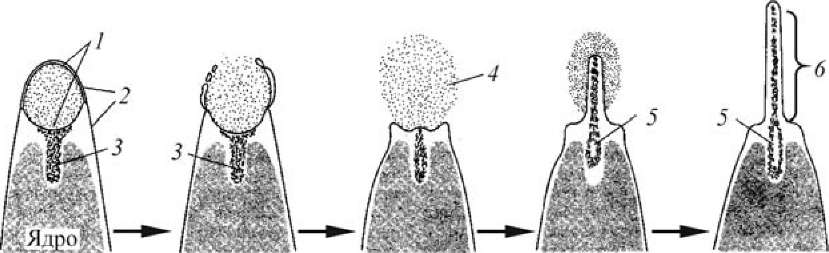 Рис. 12. Акросомная реакция спермия морского ежа (по Гилберту, 1993): 1 — акросомная мембрана; 2 — плазматическая мембрана спермия; 3 — глобулярный актин; 4 — акросомные ферменты;
5 — актиновые микрофиламенты; 6 — акросомный вырост
Рис. 12. Акросомная реакция спермия морского ежа (по Гилберту, 1993): 1 — акросомная мембрана; 2 — плазматическая мембрана спермия; 3 — глобулярный актин; 4 — акросомные ферменты;
5 — актиновые микрофиламенты; 6 — акросомный вырост
|
У морского ежа ее инициируют сульфатированные полисахариды студенистой оболочки — вызывают поступление Ca2+ в головку спермия. Это приводит к запуску процесса экзоцитоза, в результате которого акросомная мембрана сливается с цитоплазматической мембраной спермия и содержимое акросомного пузырька (прежде всего литические ферменты) выделяется в окружающую среду. Помимо этого активируется №+/Н+-обменник, что приводит к снижению внутриклеточной концентрации протонов.
В результате повышения внутриклеточного pH наблюдается усиление полимеризации глобулярного актина и формирование актиновых филамен- тов, т. е. образование акросомного выроста. Помимо этого высокий уровень pH; активирует динеиновую АТФ-азу в шейке спермия, что приводит к увеличению подвижности спермия. Однако удлинение акросомального выроста связано не только с полимеризацией актина. Приток внутрь клетки ионов кальция, натрия и хлора повышает количество осмотически активных молекул в головке спермия, что приводит к притоку в него воды. Вызванное этим резкое повышение гидростатического давления, вероятно, также способствует удлинению акросомального выроста.
Второй этап контактного взаимодействия гамет связан с узнаванием спермия и яйца.
На поверхности формирующегося акросомного выроста располагается белок (биндин), ответственный за видоспецифичное узнавание у морских ежей. На желточной оболочке яйца находится гликопротеиновый комплекс, способный образовывать связи с биндином. Таким образом, узнавание гамет у морского ежа происходит на двух уровнях: активации акросом- ной реакции и прикрепления спермия к желточной оболочке.
Контактные взаимодействия гамет у млекопитающих имеют ряд особенностей, связанных с наличием у них внутреннего оплодотворения. При этом половые пути самки принимают активное участие в процессе оплодотворения.
Спермии млекопитающих сразу после эякуляции не способны к акро- сомной реакции, для этого они должны какое-то время находиться в половых путях самки. Так, лизин, содержащийся в сперматозоидах всех млекопитающих и разрушающий прозрачную оболочку (так называемый акро- лизин), активируется только под действием гликопротеина из половых путей самки. Условия, требующиеся для капацитации (приобретение спер- мием оплодотворяющей способности), варьируют в зависимости от вида.
Природа процесса капацитации заключается:
• в изменении структуры клеточной мембраны;
• удалении с поверхности спермия особых факторов, так называемых coating factors, которые, оставаясь на поверхности, препятствуют оплодотворению.
Перестройки клеточной мембраны связаны с изменением соотношения холестерин : фосфолипиды. Снижение этого соотношения в ходе капацитации обусловлено уменьшением содержания холестерина — молекулы альбумина, имеющиеся в половых путях самки, способны отнимать холестерин у спермия. В результате происходит дестабилизация мембраны акросомного пузырька и как следствие возникает возможность осуществления акросомной реакции.
Поверхность спермия содержит фермент гликозилтрансферазу, способную узнавать концевые остатки N-ацетилглюкозамина на прозрачной оболочке яйцеклетки (рис. 13).
В спермиях, не прошедших капацитации, активные центры этого фермента блокированы связанными с их поверхностью углеводами, включающими остатки N-ацетилглюкозамина (NAr) и галактозы (Гал). При капацитации эти углеводы отделяются от поверхности спермия, освобождая активные центры галактозилтрансфераз.
Теперь галактозилтрансферазы могут узнавать N-ацетилглюкозамино- вые остатки в молекуле гликопротеина, расположенного на поверхности прозрачной оболочки и представляющего, по сути, рецептор спермия.
У млекопитающих активация сперматозоида не сопровождается ни образованием микроворсинок, ни образованием акросомного выроста. У них акросомная реакция заключается в диссоциации наружной мембраны головки сперматозоида и мембраны акросомы, но не на апикальной поверх-
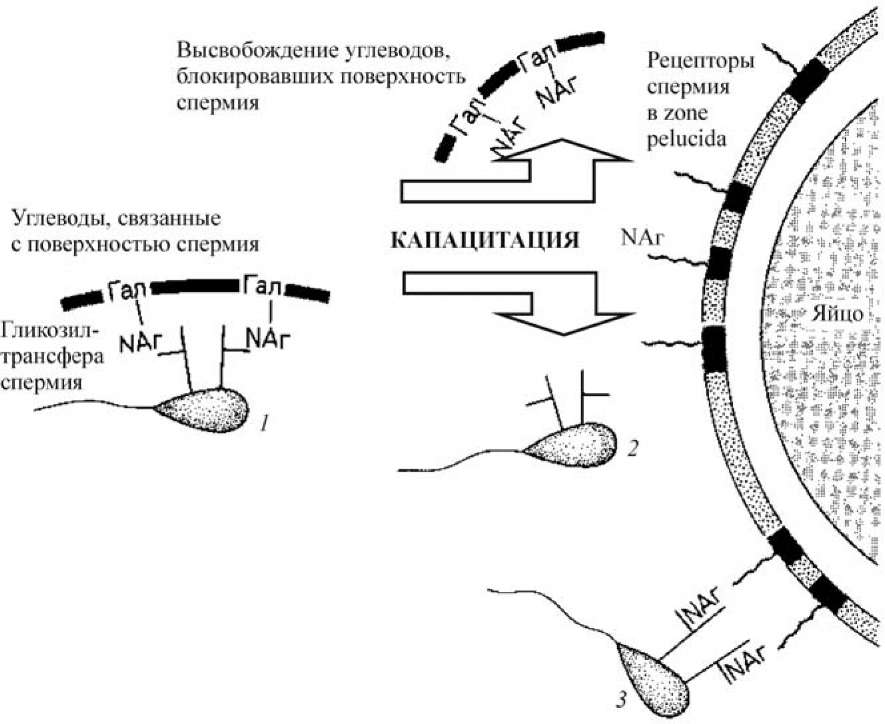 Рис. 13. Модель узнавания спермием и яйцом друг друга у млекопитающих (модель капацитации) (по Гилберту, 1993): 1 — спермий, не прошедший капацитации; 2 — капацитированный спермий со свободной гликозилтрансферазой; 3 — капацитированный спермий, прикрепляющийся к NA^-остаткам гликопротеина zona pellucida
Рис. 13. Модель узнавания спермием и яйцом друг друга у млекопитающих (модель капацитации) (по Гилберту, 1993): 1 — спермий, не прошедший капацитации; 2 — капацитированный спермий со свободной гликозилтрансферазой; 3 — капацитированный спермий, прикрепляющийся к NA^-остаткам гликопротеина zona pellucida
|
ности, а на поверхности, расположенной вдоль головки сперматозоида. Это связано с тем, что сперматозоид контактирует с яйцом не вершиной, а боком. Далее ферменты акросомы растворяют клетки лучистого венца, после чего сперматозоид вступает в контакт с блестящей оболочкой.
Блестящая оболочка яйцеклетки образована тремя типами белков: ZP1, ZP2 и ZP3. Последние два белка расположены параллельно поверхности яйца, а ZP1 сшивает эти белки, находясь перпендикулярно по отношению к ZP2 и ZP3. Контакт с ZP3 осуществляется при взаимодействии с тремя типами рецепторов спермия: терминальной галактозой, N-ацетилглюкозами- ном, гликопротеином плазматической мембраны и составляет первую часть акросомной реакции. Затем происходит выделение проакрозина (протеоли- тический фермент акросомы), который, взаимодействуя с ZP2, лизирует блестящую оболочку.
Таким образом достигается контакт задней мембраны акросомы и мембраны яйца. Молекулы, участвующие в процессе распознавания у млекопитающих, локализованы в плазматической мембране спермия, тогда как у морских ежей они располагаются в мембране акросомы. Заканчивается реакция активации сперматозоида слипанием задней мембраны акросомы спермия и мембраны яйцеклетки, их разрывом и соединением свободных концов.
В результате у яйца и сперматозоида формируется единая наружная мембрана, ограничивающая канал, через который ядро и проксимальная центриоль сперматозоида проникают в цитоплазму яйцеклетки.
Проникновение сперматозоида в яйцеклетку
Вслед за узнаванием спермия желточной (прозрачной) оболочкой яйца происходит лизис части этой оболочки в области головки спермия, в результате чего плазматические мембраны спермия и яйца сливаются. Поверхность яйца покрыта микроворсинками, контакт между сперматозоидом и яйцом вызывает полимеризацию актина и увеличение размеров микроворсинок, которые формируют воспринимающий бугорок.
Временно существующий воспринимающий бугорок в значительной степени гомологичен акросомному выросту. У морских ежей воспринимающий бугорок может возникать в любой области яйца (рис. 14).
У некоторых амфибий и многих беспозвоночных узнавание спермия и слияние с ним происходит в специализированных областях. Слияние представляет собой активный процесс, часто опосредуемый специфическими «фузогенными» (fusogenic) белками. Известно, что такие белки, как НА-бе- Л Б
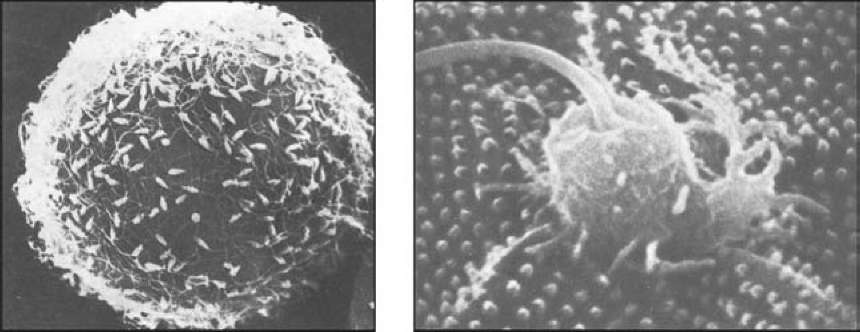 Рис. 14. Оплодотворение у морского ежа (по Гилберту, 1993): А — спермии, прикрепленные к желточной оболочке яйца; Б — спермий вовлекается в яйцо
Рис. 14. Оплодотворение у морского ежа (по Гилберту, 1993): А — спермии, прикрепленные к желточной оболочке яйца; Б — спермий вовлекается в яйцо
|
лок вируса гриппа и F-белок вируса Сендай, способствуют слиянию клеток. Не исключено, что биндин относится к белкам именно такого типа.
Предотвращение полиспермии. При нормальном моноспермном оплодотворении гаплоидные ядра спермия и яйца соединяются, образуя диплоидное ядро зиготы, при этом восстанавливается присущее виду число хромосом. Полиспермия (проникновение множества спермиев) приводит у большинства животных к гибельным последствиям. Например, у морских ежей при оплодотворении двумя спермиями возникает триплоидное ядро, в котором каждая хромосома представлена тремя, а не двумя копиями. Вместо нормального процесса — разделения хромосом с помощью биполярного веретена между двумя дочерними клетками — происходит распределение триплоидного набора между четырьмя клетками. В результате одни клетки получают лишние копии некоторых хромосом, тогда как у других эти хромосомы отсутствуют. Такие клетки либо погибают, либо развиваются атипично.
Существуют механизмы защиты яйцеклетки от полиспермии.
Быстрый блок полиспермии. Сразу после контакта первого спермия с плазматической мембраной яйца она (мембрана) должна утрачивать способность сливаться с плазматической мембраной спермия. Быстрый блок полиспермии достигает этой цели путем изменения электрического потенциала плазматической мембраны яйца (рис. 15).
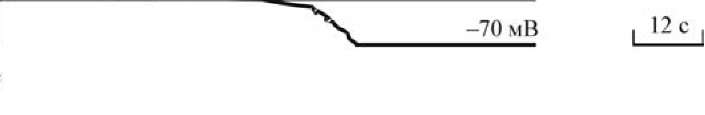 Рис. 15. Мембранный потенциал в яйце морского ежа до и после оплодотворения (по Гилберту, 1993)
Рис. 15. Мембранный потенциал в яйце морского ежа до и после оплодотворения (по Гилберту, 1993)
|
| ^ Добавление сперм ы |
| В нормальном состоянии на мембране ооцита поддерживается потенциал, равный -70 мВ. В течение 0,1 с после прикрепления первого спермия мембрана яйца деполяризуется, при этом мембранный потенциал достигает положительных величин (+20 мВ). Это является следствием резкого изменения Na+ проницаемости мембраны, в результате чего положительно заряженные ионы натрия поступают внутрь яйцеклетки (по градиенту концентрации), «уничтожая» отрицательный заряд внутренней стороны цитоплаз- матической мембраны, тем самым уменьшая исходную (-70 мВ) разность потенциалов. Открытие натриевых каналов в яйце индуцируется, по-видимому, прикреплением к нему спермия. В настоящее время из спермиев морского ежа выделен акросомный белок, способный открывать натриевые каналы в не- оплодотворенном яйце. |
____________________ ОмВ
Медленный блок полиспермии. Быстрый блок полиспермии действует недолго. Мембранный потенциал яйца морского ежа остается положительным только около 1 мин. Это кратковременное смещение потенциала может быть недостаточно для предотвращения полиспермии. Удаление избыточных спермиев, прикрепленных к желточной оболочке, осуществляется посредством кортикальной реакции. Этот более медленный блок полиспермии начинает действовать примерно через 1 мин после прикрепления спермия к яйцу.
В яйце морского ежа непосредственно под плазматической мембраной располагаются около 15 000 кортикальных гранул. После контакта спермия с яйцом в присутствии Ca2+ эти кортикальные гранулы сливаются с плазматической мембраной и выделяют содержимое в область между плазматической мембраной и желточной оболочкой (рис. 16).
Белки, связывающие желточную оболочку и поверхность яйца, разрушаются высвобождающимися пептидазами:
• вителлиновая деламиназа — отделяет желточную оболочку от цито- плазматической мембраны яйцеклетки;
• сперморецепторная гидролаза — освобождает поверхность яйца от осевших на желточной оболочке сперматозоидов, лизируя сайты их соединения.
Выделяемые мукополисахариды и осмотически активный гликопроте- ид создают осмотический градиент, обусловливающий поступление воды из цитоплазмы яйца в пространство между желточной оболочкой и плазматической мембраной (перивителлиновое пространство). В результате объем яйцеклетки несколько уменьшается, желточная оболочка отделяется от поверхности яйца и с этого момента называется оболочкой оплодотворения. В процессе кортикальной реакции она претерпевает ряд изменений:
• воздействие протеаз приводит к изменению свойств биндиновых рецепторов или к их отделению вместе с прикрепленными к ним спермиями;
Ферменты из кортикальных гранул разрушают связи желточной оболочки
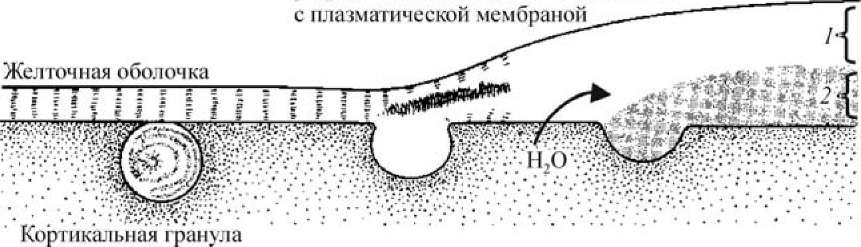 Рис. 16. Формирование оболочки оплодотворения (по Гилберту, 1993): 1 — оболочка оплодотворения; 2 — гиалиновый слой
Рис. 16. Формирование оболочки оплодотворения (по Гилберту, 1993): 1 — оболочка оплодотворения; 2 — гиалиновый слой
|
• пероксидаза, содержащаяся в кортикальных гранулах, вызывает затвердевание оболочки путем образования поперечных связей между остатками тирозина соседствующих белков.
Образование оболочки оплодотворения начинается в месте проникновения спермия (примерно через 20 с после прикрепления) и отсюда распространяется по всей поверхности яйца, завершаясь к концу первой минуты после прикрепления оплодотворяющего спермия. Одновременно происходит выделение гиалина — белка, запасенного в кортикальных гранулах, который образует сплошной слой вокруг яйца. Плазматическая мембрана взаимодействует с этим белком, и гиалиновый слой поддерживает бластомеры в период дробления.
У млекопитающих кортикальная реакция не вызывает образования оболочки оплодотворения, но результат ее оказывается таким же, как и у морских ежей: рецепторы спермиев изменяются, и они больше не удерживают сперматозоиды на поверхности яйца. Этот процесс изменения свойств рецепторов спермиев носит название реакции прозрачной оболочки или реакции zona.
У млекопитающих частота полиспермии бывает минимальной благодаря малому числу спермиев, достигающих места, где происходит оплодотворение. Механизмы защиты от полиспермии, наблюдаемой при оплодотворении богатых желтком яиц (некоторые птицы, рептилии, амфибии), когда в цитоплазму проникает несколько сперматозоидов, неясны. При этом все спермии, за исключением одного, разрушаются, после того как женский пронуклеус сольется с мужским.
10 Механизм кортикальной реакции сходен с механизмом акросомной реакции. Прикрепление спермия к плазматической мембране яйца через активацию находящегося в мембране G-белка стимулирует активность другого мембраносвязанного фермента — фосфолипазы С. Она, в свою очередь, расщепляет находящийся здесь же фосфатидилинозитол-4,5-бифосфат на диацилглицерол — ДАГ (остается связанным с мембраной), и инозитолтри- фосфат — ИТФ (диффундирует в цитоплазму).
Диацилглицерол посредством протеинкиназы С активирует №+/Н+-ан- типортер, что приводит к увеличению внутриклеточного pH (натрий входит в клетку, а протон ее покидает) и как следствие активации белкового синтеза, репликации ДНК и перемещения морфогенетических детерминант в цитоплазме (при участии Ca2+). ИТФ вызывает высвобождение Ca2+ из внутриклеточных депо (эндоплазматического ретикулума), что приводит к экзо- цитозу кортикальных гранул и включению медленного блока полиспермии, а также образованию гиалинового слоя вокруг яйца.
Другим Ca2+-зависимым эффектом является активация НАД-киназы, которая катализирует превращение НАД в НАДФ+, что в конечном итоге обеспечивает синтез новых липидных компонентов плазматической мембраны, необходимых для последующего протекания процесса дробления.
Слияние генетического материала
У морских ежей головка спермия проникает в яйцо перпендикулярно его поверхности. После слияния мембран сперматическое ядро и центриоль отделяются от митохондрий и жгутика, которые разрушаются и в цитоплазме развивающихся взрослых организмов не обнаруживаются.
Таким образом, митохондрии зародышу передаются только от материнского организма, а центриоли достаются от отцовского, поскольку яйцеклетка утрачивает свои центриоли при последнем делении мейоза.
Ядро яйца после завершения второго деления созревания и приобретения пузыревидной формы называется женским пронуклеусом. Сперматическое ядро в цитоплазме яйца деконденсируется и преобразуется в мужской пронуклеус.
Ядерная оболочка сперматического ядра распадается на мелкие пузырьки, что делает возможным воздействие цитоплазмы яйца на компактный хроматин спермия. После проникновения спермия в цитоплазму яйца мужской пронуклеус совершает поворот на 180о так, что центриоль спермия оказывается расположенной между мужским и женским пронуклеусом. Ее микротрубочки удлиняются, вступая в контакт с женским пронуклеусом, после чего пронуклеусы перемещаются навстречу друг другу.
У морского ежа в результате слияния пронуклеусов возникает диплоидное ядро зиготы. Синтез ДНК инициируется на стадии пронуклеусов (в период их миграции) либо после образования ядра зиготы.
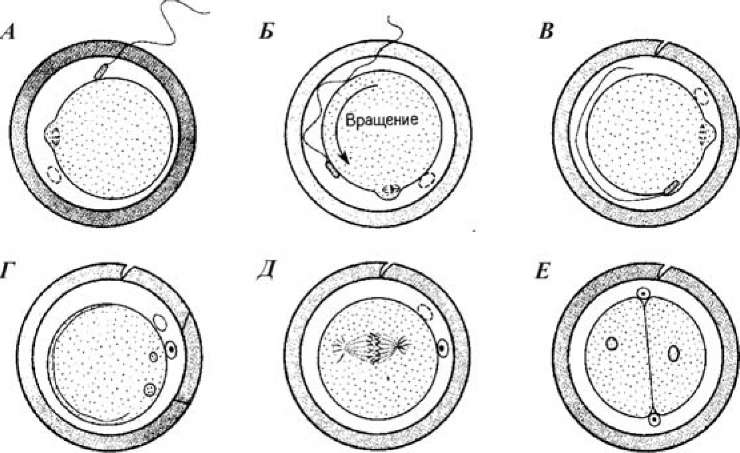 Рис. 17. Схема процесса оплодотворения у хомячка (по Гилберту, 1993): А — спермий прикрепляется к яйцу; Б — прикрепление спермия к яйцу и взмахи его хвоста вызывают вращение яйца; В — плазматические мембраны спермия и яйца сливаются; Г — формируются пронуклеусы; Д — начинается первое деление дробления; Е—двухклеточный зародыш
Рис. 17. Схема процесса оплодотворения у хомячка (по Гилберту, 1993): А — спермий прикрепляется к яйцу; Б — прикрепление спермия к яйцу и взмахи его хвоста вызывают вращение яйца; В — плазматические мембраны спермия и яйца сливаются; Г — формируются пронуклеусы; Д — начинается первое деление дробления; Е—двухклеточный зародыш
|
А Б В
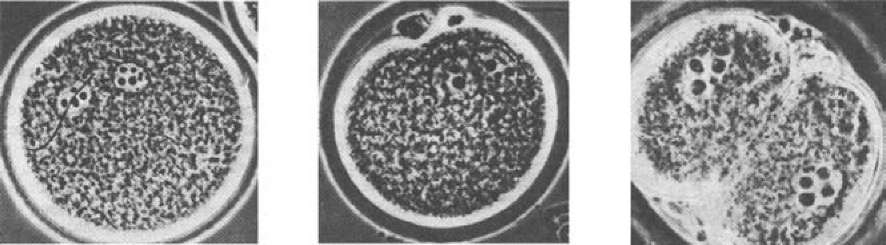 Рис. 18. Оплодотворение у хомячка (по Гилберту, 1993): А — проникновение спермия в яйцо и набухание мужского про- нуклеуса; Б — установление контакта между мужским и женским пронуклеусами; В — двухклеточная стадия
Рис. 18. Оплодотворение у хомячка (по Гилберту, 1993): А — проникновение спермия в яйцо и набухание мужского про- нуклеуса; Б — установление контакта между мужским и женским пронуклеусами; В — двухклеточная стадия
|
У млекопитающих процесс сближения пронуклеусов (рис. 17) продолжается около 12 часов (у морского ежа — всего 1 час). Головка спермия у млекопитающих проникает не перпендикулярно поверхности яйца, а почти по касательной к ней, сливаясь с многочисленными микроворсинками. Пока ядро ооцита млекопитающего завершает второе мейотическое деление, мужской пронуклеус увеличивается в размерах. Затем оба пронуклеуса перемещаются навстречу друг другу, реплицируя ДНК в процессе миграции.
Когда пронуклеусы приходят в контакт, их ядерные оболочки разрушаются (рис. 18). Происходит конденсация хроматина с образованием видимых хромосом, которые (у большинства животных) располагаются на общем митотическом веретене первого деления дробления (центриоли присутствуют в яйцеклетках млекопитающих изначально, а не передаются зародышу от спермия). Таким образом, у млекопитающих истинно диплоидное ядро впервые появляется не у зиготы, а у двухклеточного зародыша.
Преобразование цитоплазмы яйца
После проникновения сперматозоида скорость метаболизма увеличивается в 70—80 раз, начинаются интенсивные перемещения слоев цитоплазмы яйцеклеток друг относительно друга. Иногда при этом происходит расслоение различных частей ооплазмы — ооплазматическая сегрегация. В ходе этого процесса намечаются основные, хотя далеко не все, элементы пространственной организации зародыша. Поэтому данный этап называют проморфогенезом, так как именно тогда расставляются вехи для будущих морфогенетических процессов.
У некоторых кишечнополостных сегрегация ограничивается процессом разделения ооплазмы на внешний ободок эктоплазмы (содержащий различные пигменты и бедный питательными включениями) и внутреннюю массу эндоплазмы (богата желтком и другими питательными включениями). Уже такое достаточно простое расслоение влияет на последующие процессы развития, определяя в яйцах радиальную установку веретен делений дробления. Так, в яйцеклетках амфибий положение сагиттальной плоскости и проходящей в ней дорсо-вентральной оси определяется именно процессами ооплазматической сегрегации, которые протекают в первые десятки минут после оплодотворения. Реорганизации цитоплазмы зиготы способствуют микротрубочки спермия (при добавлении колхицина перемещения цитоплазматических масс не происходит). В большинстве случаев ооплазматическая сегрегация влияет не на конечную дифференцировку клеток, а на ближайшие этапы развития — дробление и гаструляцию.
11 Партеногенез — развитие, происходящее без оплодотворения (участия сперматозоида). Естественный партеногенез обнаружен у ос, пчел, ряда чешуекрылых, у некоторых видов ящериц и змей, типичен для летних поколений ряда ракообразных и коловраток. Успешные опыты по искусственному партеногенезу были выполнены А. А. Тихомировым в 1886 году. Ему удалось стимулировать развитие неоплодотворенных яиц тутового шелкопряда кратковременным нагреванием или потиранием их щеткой. Естественный партеногенез чаще всего случается при незавершенном оплодотворении, т. е. в тех случаях, когда имела место активация яйцеклетки, но ядро сперматозоида не участвовало в оплодотворении. В активированных яйцах используется генетическая информация, содержащаяся только в женском пронуклеусе (гиногенез). При искусственном партеногенезе, когда существует возможность экспериментально удалить женский пронуклеус, развитие зародыша может осуществиться только за счет мужского пронуклеуса (андрогенез). В указанных случаях потомки целиком наследуют либо признаки матери (при гиногенезе), либо отца (при андрогенезе). Это лишний раз свидетельствует о том, что наследственные свойства особи определяются в основном ядром, а не цитоплазмой. В большинстве случаев после пар- теногенетической активации яйцеклетки в ней восстанавливается диплоидный набор хромосом.
У млекопитающих также отмечались случаи вступления яйцеклеток на путь партеногенетического развития — самопроизвольно либо под влиянием активирующего агента (электростимуляция, тепловой шок, этанол). Однако развитие таких зародышей всегда останавливалось на ранних стадиях — рождения партеногенетически развившихся особей никогда не наблюдалось. Полноценное развитие партеногенетиков у млекопитающих невозможно, по-видимому, вследствие того, что в женских хромосомах заблокированы (в результате метилирования) определенные участки ДНК, присутствующие в активной форме в мужских хромосомах. Именно поэтому самец не может быть заменен партеногенетическим агентом. Экспериментально показано, что если у мыши при мейозе подавить отделение полярного тельца, то возникнут и начнут развиваться диплоидные яйцеклетки, в которых весь наследственный материал берет начало от материнского организма. Такие клетки делятся и образуют зародышей с хордой, мышцами, скелетом и разными органами, включая бьющееся сердце. Но развитие продолжается недолго — уже на 10—11-е сутки между нормальными и парте- ногенетическими зародышами обнаруживаются глубокие различия.
Таким образом, хотя у многих животных оба пронуклеуса эквивалентны, у млекопитающих между ними имеются важные функциональные различия. Для завершения развития млекопитающих необходимы оба пронуклеуса: материнский и отцовский.
Дата добавления: 2016-12-27; просмотров: 6179;
Поиск по сайту
Узнать еще
- Cравнительная характеристика усилителей на БТ
- I. Загальна характеристика
- II. Монозы и их характеристика
- Nemathelmintes. Общая характеристика типа. Nematoda. Характеристика класса. Медицинское значение. Био- и геогельминты.
- Plahelmintes (Плоские черви).Общая характеристика типа. Морфология, систематика, основные представители, значение.
- Protozoa. Общая характеристика подцарства. Классификация. Представители. Медицинское значение.
- V. Механическая характеристика
- V. Упрощенная схема замещения трансформатора и внешняя характеристика.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
