Место популяционной экологии в системе экологических дисциплин
Изучение биосферы возможно только с использованием системного подхода. Это определяется естественной дифференциацией всего живого на соподчиненные уровни. В соответствии с этой дифференциацией сложилась и структура биологических дисциплин.Так, все вы знаете, что исследования в области биохимии и молекулярной биологии выполняются на молекулярном уровне, цитология изучает строение и функции клеток и их структур, функционирование и строение организмов являются предметом изучения анатомии и физиологии животных и растений. И так далее – список можно продолжить. В то же время правомерно утверждать, что познанием наиболее общих закономерностей взаимосвязей всего живого с окружающей средой занимается экология.
Экологические исследования выполняются на разных уровнях: организменном, популяционном, на уровне сообществ и на уровне целых экосистем. Перечисленным уровням соответствуют частные экологические дисциплины, а именно аутэкология, популяционная экология, синэкология (=экология сообществ) и экология экосистем. Популяционная экология, основы которой вам предстоит освоить, является частью более общей дисциплины – популяционной биологии. Частью популяционной биологии является также популяционная генетика – наука, занимающаяся изучением частот генотипов и микроэволюционными процессами в популяциях.
Основным предметом популяционной экологии является структура и динамика популяций, а также определяющие их факторы. В связи с этим основные задачи, или вопросы, на которые отвечает популяционная экология, можно сформулировать так:
1) почему те или иные организмы в данный момент времени встречаются в этом, а не в каком-либо другом месте?
2) почему численность или биомасса данных организмов именно такая, а не какая-либо другая?
3) если численность и биомасса особей меняется во времени, то почему именно так, а не как-нибудь по-другому?
Ответы на эти вопросы важны не только с теоретических, но и с практических соображений. Например, они позволяют соответствующим образом планировать использование популяций диких и домашних видов, а также осуществлять мероприятия по охране исчезающих видов.
1.2. Определение популяции
Оказывается, что дать определение понятию популяции не так уж легко. Причин этому несколько.
1) Популяция как объект исследования интересует не только экологов, но, как мы уже сказали, и генетиков, а также специалистов по микроэволюции. Проблема здесь в том, что разные специалисты применяют разные подходы при определении популяции.
2) Популяции часто имеют сложную иерархическую структуру, т.е. подразделяются на более мелкие структурные единицы (например, метапопуляции, парцеллы, демы, семьи и т.д.). Разногласия здесь появляются из-за того, что разные исследователи придают разную значимость этим уровням иерархии.
3) Кроме того, понятно, что организмы, принадлежащие к разным систематическим группам, представляют собой очень разные объекты, которые только условно можно обозначить словом популяция. Например, микробиолог под термином «популяция» в отношении к какой-либо бактерии подразумевает совсем не то, что ботаник, который изучает луговую растительность или зоолог, изучающий слонов.
Еще в Древних Греции и Риме словом «популяция» обозначали человеческое общество или собрание, группу людей. В качестве научного термина это понятие впервые было использовано в 1903 г. Иогансеном. Впоследствии многие исследователи давали свои определения «популяции». Приведем несколько из них.
Начнем с того, что очень часто экологи вообще не задумываются над определением популяции, а пользуются этим термином для обозначения любой совокупности особей одного вида, населяющих какую-то более или менее однородную территорию (или акваторию) или содержащихся в лабораторных условиях. Такая точка зрения фактически отражена в определении популяции, данном Перлом (Pearl, 1937) еще в 1930-х гг.: «Популяция– группа живых особей, выделяемая в некоторых рамках пространства и времени».
«Популяция – совокупность особей одного вида, обитающих на определенной территории» (Гиляров, 1981).
«Популяция – совокупность особей вида в определенном биотопе» (Хадорн, Венер, 1989).
«Популяция – группа особей определенного вида, вероятность скрещивания которых друг с другом велика по сравнению с вероятностью скрещивания с представителями какой-то другой группы» (Пианка, 1981).
Есть и такое длинное определение: «Популяция – совокупность особей одного вида, в течение достаточно длительного времени населяющих определенное пространство, внутри которого практически осуществляется определенная степень панмиксии и нет заметных изоляционных барьеров, которая отделена от соседних таких же совокупностей особей данного вида определенной степенью давления определенных форм изоляции» (Тимофеев-Ресовский, Яблоков, Глотов, 1973).
Одним из наиболее удачных определений, соответствующих современным представлениям о популяциях, пожалуй, является определение, данное Яблоковым (1987): «Популяция – минимальная самовоспроизводящаяся группа особей одного вида, на протяжении эволюционно длительного времени населяющая определенное пространство, образующая самостоятельную генетическую систему и формирующая собственное экологическое гиперпространство» (Яблоков, 1987).
Некоторые из положений этого определения заслуживают специального внимания:
1) Минимально самовоспроизводящаяся группа – группа особей, которая может участвовать в эволюционных процессах. При резких изменениях факторов среды популяция для длительного выживания должна обладать определенной минимальной численностью.
2) Самостоятельная генетическая система. Если ввести в понятие «популяция» критерий скрещиваемости (панмиксии), то он исключит из сферы применения этого понятия группу агамных и облигатно партеногенетических видов. Агамия – размножение без участия половых клеток, т.е. деление, почкование. Агамия свойственная многим растениям и некоторым беспозвоночным. Партеногенез –размножение из женской половой клетки без участия мужской клетки. Так вот: несмотря на указанные способы размножения, и у агамных, и у партеногенетических форм популяции состоят из генетически разнородных особей, т.е. формируют самостоятельные генетические системы. Нередко агамные популяции генетически и фенотипически оказываются даже более изменчивыми, чем популяции с половым размножением.
3) Собственное экологическое гиперпространство.С концепцией экологической ниши и гиперпространства мы познакомимся более подробно позднее. Сейчас же отметим, что для каждой популяции одного и того же вида свойственны свои экологические характеристики. В связи с этим ареал популяции может расширяться только при условии, что популяция сможет освоить новое пространство и его ресурсы.
Понятно, что идеального в нашем мире ничего нет. Это в равной степени касается и определений популяции. Например, практически все они отмечают, что особи одной популяции населяют определенное пространство. Но как, например, быть с гетеротопными видами, т.е. животными или растениями, у которых разные стадии жизненного цикла обитают в разных средах? Примером здесь могут быть комары, личинки которых живут в водоемах, а по завершении метаморфоза вылетают и становятся наземными животными. Другой пример – паразитические организмы, например, трематоды. Обычно после партеногенетического размножения в первом промежуточном хозяине они выходят в воду в виде планктонных личинок церкарий. Обычно, задача церкарий состоит в том, чтобы отыскать второго промежуточного хозяина. Пока же церкарии находятся в составе планктона, они фактически представляют собой самостоятельную популяцию.
В отношении названных организмов В. Н. Беклемишев предложил термин гемипопуляция. Гемипопуляция (hemipopulation) – это самостоятельная часть популяции, представленная определенной стадией жизненного цикла организма.
В случае с паразитическими организмами одним термином «гемипопуляция» не обойтись. Дело здесь обстоит еще сложнее, и для паразитов выделяют еще ряд популяционных группировок, например:
1) Макропопуляция (component population) – совокупность всех гемипопуляций одного вида паразита в популяции хозяина.
2) Микропопуляция (infrapopulation) – совокупность всех особей одного вида паразита внутри или на одной особи хозяина.
Вывод здесь один – эколог, по возможности, должен пристально наблюдать за всеми стадиями жизненного цикла изучаемого вида, вне зависимости от того, для каких экосистем или биотопов эти стадии традиционно указываются. Только в этом случае можно понять те основные процессы, которые происходят в исследуемой популяции.
В этой связи можно привести еще один пример – с морской звездой терновый венец Acanthaster planci. В тропических районах Индийского и Тихого океанов численность этого животного периодически проявляет вспышки. Это сопровождается опустошением коралловых рифов, поскольку терновый венец питается мягкими тканями кораллов. Причины вспышек численности животного искали и в хищничестве, и в конкуренции, и в других механизмах. Но оказалось, что все дело в повышении выживаемости планктонных личинок звезды, вызванном периодическим повышением численности фитопланктона, которым личинки питаются. Вспышки же фитопланктона были обусловлены смывом биогенов с прибрежных территорий.
1.3. Пространственно-временной масштаб изучения популяций
При изучении популяций и выяснении их места в экосистемах необходимо также представлять себе, что каждая популяция существует как бы в своем пространстве и времени. Поясним это на следующем простом примере.
Одной из наиболее обычных тлей в зоне Евразии является Cinaria pini. Это насекомое может всю свою жизнь провести на одной хвоинке сосны, высасывая соки из ее тканей. Популяция тлей в течение ряда лет может существовать на одном дереве. Кормовое растение с его микроклиматом и населением является целой экосистемой для популяции тлей. То «определенное пространство», которое упоминается в определениях популяции, с «точки зрения» тлей представлено одним деревом и его непосредственным окружением.
Далее. Предположим, что сосна, на которой обитают тли, растет в смешанном лесу. В таком лесу самое обычное млекопитающее – это лесная полевка Clethrionomys glareolus. Жизнь одной полевки обычно протекает на участке в несколько сотен или тысяч квадратных метров. Популяция же полевок занимает пространство в десятки, и даже сотни квадратных километров. Экосистема с точки зрения полевки – это уже целый участок леса со своим микроклиматом, растительностью, хищниками и конкурентами.
Если мы теперь обратимся к самому крупному млекопитающему в том же лесу – лося Alces alces, то столкнемся уже с совершенно другими масштабами пространства. За сутки лось может проходить несколько километров. Поэтому популяция лося занимает сотни и даже тысячи квадратных километров.
Поскольку популяции тлей, полевок и лосей занимают такие разные площади, изучение их, очевидно, должно требовать разного пространственного масштаба.
Аналогичные рассуждения применимы и в отношении временного масштаба изучения рассмотренных популяций. Это обусловлено разными скоростями протекающих в них процессов.
Известно, например, что плодовитость одной самки тли за один летний сезон может составить более тысячи новых особей. Численность же тлей в течение года может колебаться в 100-1000 раз. Пара полевок за весенне-летний сезон в удачный год может дать 30-40 особей приплода. Численность же популяции полевок за год может изменяться лишь в десятки раз. Наконец, лоси размножаются не чаще чем раз в год, и самка приносит лишь 1-2 детенышей. При этом для достижения половой зрелости самке лося необходимо 3-4 года. Следовательно, значимые изменения численности лосей происходят в течение десятков лет.
Таким образом, очевидно, что изучение механизмов популяционной динамики тлей требует очень частого (по крайней мере, еженедельного) обследования популяции, а период наблюдений должен охватывать год-два. Аналогичные наблюдения за полевками должны длиться уже 5-10 лет. При этом сами обследования популяции будут проводиться, например, раз в несколько месяцев. Наконец, в случае с лосями изучение популяционной динамики потребует десятков лет.
1.4. Эмерджентные свойства популяций
Любая популяция обладает рядом характеризующих ее признаков:
1) статические:численность (плотность), характер пространственного распределения, возрастная и генетическая структура.
2) динамические: рождаемость и смертность, динамика численности.
Таким образом, популяции обладают как биологическими, так и групповыми свойствами. Что это означает?
1) Биологические свойства популяции определяются свойствами входящих в нее особей. К таким свойствам относят наличие жизненного цикла у популяции, ее способность к росту, дифференцировке и самоподдержанию.
2) Поскольку популяция – это совокупность особей, она обладает и групповыми характеристиками, а именно: рождаемость, смертность, размерно-возрастная структура, генетическая пластичность. Эти признаки относятся только к популяции. Почему?
Особь рождается и умирает, имеет тот или иной возраст, но применительно к особи нельзя говорить о рождаемости, смертности, размерной или возрастной структуре. Эти свойства, отсутствующие у особей, но возникающие у группы, т.е. отличающие один уровень организации от другого, называют эмерджентнымисвойствами (от англ. emergent – внезапно возникающий).
Рассмотрим более подробно черты сходства и различий между популяцией и составляющими ее особями. В отличиеот организма, популяция:
· не имеет морфологической отграниченности от среды и других популяций;
· в популяциях не вычленяются части, аналогичные физиологически функциональным системам и органам организма;
· организм как биологическая система относительно недолговечен, популяция же при сохранении необходимых условий практически бессмертна.
Есть и черты сходства популяции с организмом как системой, которые позволяют популяцию также квалифицировать как биологическую систему:
· структурированность пространственная и функциональная – особи популяции обычно распределяются не хаотично, а определенным образом, т.е. занимают то «определенное пространство» о котором говорится практически в любом определении популяции. Функции особей в популяции обычно тоже различаются (самки и самцы, например).
· интегрированность составных частей – особи в популяции непрерывно обмениваются информацией, и это общение представляет собой специфический механизм формирования и поддержания целостности популяции как единой системы.
· саморегуляция – обычно при сдвигах в функционировании популяции под действием внутренних или внешних условий, она, как и организм, стремится вернуться к исходному состоянию или адаптироваться к новым условиям.
В связи с тем, что популяции обладают свойствами и организмов, и группы, некоторые исследователи впадают в крайности относительно методологии изучения популяций.
Одной из крайностей является органицизм – это такой подход, при котором популяция считается в высшей степени целостном образованием, реагирующее на любое изменение окружающей среды так, как реагировал бы отдельный организм. Принимается, что «стратегическая задача» каждой популяции заключается в поддержании стабильного существования путем занимания определенной территории и сохранения определенной численности.
Другая крайность – это редукционизм: сведение поведения сложного объекта к поведению его взаимодействующих элементов. Редукционисты убеждены, что популяция как целое вообще не представляет никакой реальности, т.е. является воображаемым идеальным объектом, а так называемое «поведение популяций» (т.е. целесообразное реагирование на изменение среды) есть не что иное, как просто удобный для исследователей способ описаниярезультатов суммарной активности многих отдельных особей.
1.5. Популяционная экология и математическое моделирование
Вам сразу необходимо «смириться с мыслью», что популяционная экология – это одна из самых формализованных областей биологии. Это означает, что в популяционной экологии сплошь и рядом используются методы математики и статистики. Применение этих методов обусловлено потребностью создавать модели популяционных процессов для прогнозирования развития популяций. Модель – упрощенный аналог реального объекта, явления и процесса.Фактически модель представляет собой математическое описание того или иного процесса в популяции. Она может быть представлена простым уравнением с одной переменной или сложной системой с сотнями уравнений, содержащими десятки переменных.
Самое сложное при построении модели – это решение о том, насколько она должна быть сложна в каждой конкретной ситуации. Ответ на этот вопрос определяется тремя факторами:
1) свойствами изучаемого вида (его экологией);
2) объемом знаний об этом виде (т.е. доступность необходимых данных)
3) вопросом, на который должна отвечать модель.
Модель и реальный мир связаны между собой двумя процессами: абстракцией и интерпретацией. Абстракция означает обобщение, т.е. выделение наиболее важных компонентов реальных систем и пренебрежение менее важными компонентами. Важность компонентов определяется исходя из их роли в динамике системы, в нашем случае – в динамике популяции. Например, если мы установили, что изучаемая популяция заражена паразитами лишь на 5%, мы можем смело исключать фактор паразитизма из модели.
Интерпретация –это соотнесение результатов моделирования с реальной ситуацией.Если компоненты модели (т.е. параметры и переменные), а также ее поведение неправильно отражают компоненты, характеристики и поведение реальной системы, то модель бесполезна.
Большинство экологов, работающих в полевых условиях, не очень сильны в осуществлении абстракции. При построении модели, они пытаются включить в нее все имеющиеся детали. В то же время большинство математиков не сильны в интерпретации построенных ими моделей. Однако и абстракция, и интерпретация одинаково важны при моделировании. Поэтому очень важным в наши дни становится тесное сотрудничество между экологами и математиками.
2. ЧИСЛЕННОСТЬ И ПЛОТНОСТЬ ПОПУЛЯЦИИ И МЕТОДЫ ИХ ОЦЕНКИ
2.1. Соотношение понятий «плотность» и «численность» популяции
Следует отметить, что в русскоязычной научной литературе понятие «плотность популяции» часто отождествляют с понятием «численность популяции». Тем не менее, между этим термины следует разграничивать:
1) Общая численность популяции – это общее количество особей, которые образуют популяцию. В англоязычной литературе этому термину соответствует «total number/size of population».
2) Плотность популяции – это количество особей в расчете на единицу пространства или объема (м2, м3). В англоязычных публикациях практически всегда этому понятию соответствует «density of population», реже «abundance of population».
Некоторые особенности терминологии имеются в случае паразитами и их популяциями и внутрипопуляционными группировками. Так, количество особей паразита с простым жизненным циклом в одной особи хозяина называется численностью микропопуляции.Этот термин является синонимом широко употребляемого в паразитологии термина интенсивность инвазии (заражения) (экз./особь). Очевидно, что для оценки всей численности таких паразитов, например, в целом по водоему, необходимо умножить среднюю численность микропопуляций на общую численность хозяина в водоеме. Однако в большинстве случаев паразиты обладают сложным жизненным циклом, т.е. со сменой хозяев и сред обитания. Если помните из первой лекции, мы говорили о том, что совокупность всех особей паразита, находящихся на определенной стадии развития, называется гемипопуляцией. Таким образом, у паразитов можно изучать численности отдельных гемипопуляций. Так, например, совокупность всех особей паразита, находящихся на определенной стадии развития, в одной особи хозяина называется численностью микрогемипопуляции.Задача нахождения общей численности гемипопуляций у паразитов со сложным жизненным циклом значительно усложняется, если в качестве хозяев паразит может использовать несколько разных видов хозяев. В таких случаях для нахождения общей численности определенной гемипопуляции необходимо оценить численность популяций уже всех возможных хозяев.
2.2. Методы определения общей численности популяций
Необходимость определения общей численности популяций возникает не часто. Исключение составляют случаи, когда необходимо выяснить, например, общую численность ли поголовье видов, важных с хозяйственной точки зрения. Подобные оценки важны также при определении общей численности видов, находящихся под угрозой исчезновения, например, животных занесенных в Красную книгу. Еще одна область применения таких подсчетов – это выяснение количества тех или иных видов в национальных парках, зоопарках, заповедниках и т.д.
Очевидно, что определить численность всех особей, образующих популяцию крайне сложно. Но иногда это возможно, если речь идет о крупных и хорошо заметных для человека организмах, образующих немногочисленные скопления на ограниченной территории. Например, дикие северные олени Кольского полуострова в конце зимы - начале весны скапливаются на небольших горных возвышенностях, где могут докопаться до лишайников, служащих им кормом. Аналогично, по «головам» можно сосчитать и колониально гнездящихся птиц – кайр, грачей, белых гусей и т.д. Для определения численности подобных видов часто используют фотографирование, которое исследователь выполняет с самолета или вертолета. Впоследствии по фотографиям можно легко подсчитать всех животных. Когда-то такой способ был использован немецким зоологом Бернардом Гржимеком для оценки численности различных африканских копытных в африканском национальном парке «Серенгети».
Для оценки общей численности относительно крупных и подвижных животных часто используется метод мечения с последующим отловом (capture-recapture method). Этот метод был предложен известным гидробиологом – Петерсеном. Его суть в том, что животных ловят, определенным образом метят и выпускают обратно в природу, туда, где они были пойманы. Через некоторое время производят новый облов, и по доле, которую составляют меченые особи от общего числа пойманных, определяют численность популяции:
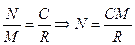 , где
, где
М – количество особей, меченых в первой выборке;
С – количество особей, отловленных во второй раз;
R – количество меченых особей во второй выборке.
Было показано, что в случае с небольшими выборками данная формула дает завышенные результаты. Поэтому в нее вводят поправку:
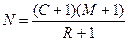
Для того чтобы данный метод «работал» корректно, необходимо выполнение нескольких условий:
1) Популяция является закрытой. Т.е. в ней не наблюдаются процессы иммиграции и эмиграции особей.
2) Выловленные и меченые животные должны представлять случайную выборку из популяции, т.е. в этой выборке не должен быть повышен процент слабых, больных или малоактивных (слишком активных) животных.
3) Выпущенные в природу меченые животные должны полностью перемешаться с остальной частью популяции.
4) Вероятность выживания меченых особей должна быть примерно такой же, как немеченых особей.
5) При вторичном отлове вероятность поимки меченых животных не должна быть больше или меньше, чем немеченых особей.
6) Время между двумя отловами должно быть меньше продолжительности жизни одного поколения.
Перечисленные условия на практике редко выполняются полностью, однако метод мечения довольно широко применяется, например, при оценке численности рыб, обитающих в замкнутых водоемах. Иногда он оказывается единственно возможным для таких целей.
Рассмотрим еще два метода, позволяющих оценить общую численность животных. Первый – это так называемый метод Келкера (Kelker 1940), предложенный для промысловых видов. Метод применяется в случаях, когда в популяции избирательно элиминируются особи с определенным признаком, отсутствующим у другой части этой популяции. Такие особи, различающиеся по какому-то признаку, например, по полу, принято обозначать как организмы х-типа и у-типа. Чтобы разобраться с методом Келкера, рассмотрим гипотетическую популяцию, в которой преимущественно отстреливают самцов. Очевидно, что соотношение полов в популяции до проведения охоты и после будет разным. При этом доля самцов в популяции после проведения охоты будет определяться следующим образом:
| {Доля самцов после охоты} = | {Число самцов до охоты} – {Число убитых самцов} |
| {Численность популяции до охоты} – {Число убитых животных} |
(S)Введем следующие обозначения:
N1 – общая численность популяции до охоты;
N2 – общая численность популяции после охоты;
Х1 и Х2 – число самцов в популяции до и после охоты;
Y1 и Y2 – число самок в популяции до и после охоты;
р1 = Х1/N1 – доля самцов в популяции до охоты;
р2 = Х2/N2 – доля самок в популяции до охоты;
Rx = X2 - X1 – изменение численности самцов за период проведение охоты;
Ry = Y2 - Y1 – изменение численности самок за период проведение охоты;
R = Rx - Ry – изменение общей численности популяции.
С учетом этих обозначений модель Келкера в формализованном виде можно записать так:
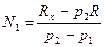
Давайте рассмотрим, как применять эту формулу на таком примере. До проведения сезона охоты сотрудниками заказника были отловлены 1400 фазанов. Среди них 600 птиц оказались самцами. После проведения охоты провели повторное обследование популяции и выяснили, что из 2000 отловленных птиц самцами были только 200. Всего за сезон были убиты 8000 самцов и 500 самок. Необходимо оценить численность популяции до проведения охоты.
Находим необходимые параметры:
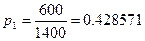
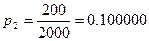
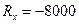
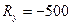
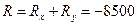
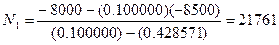
При этом после охоты численность популяции птиц будет составлять:
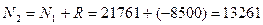
Наконец, еще один метод определения общей численности популяций – это метод Лесли и Девиса (Leslie, Davis 1939). Он применяется в случае с популяциями, используемыми в промысловых целях. Для таких популяций характерно изъятие значительно числа особей с каждым последующим промысловым усилием. В качестве промыслового усилия здесь имеется в виду, например, одноразовое забрасывание невода, отлов организмов вдоль одной трансекты и т.д. Применение метода требует соблюдение следующих трех условий:
1) Популяция является закрытой.
2) Поимка одной особи никак не влияет на вероятность поимки другой;
3) Вероятность попадания в улов постоянна для всех особей популяции.
Кроме того, в каждом из последовательных отловов (отстрелов) должно применяется одно и то же промысловое усилие.
Суть метода Лесли и Дэвиса рассмотри на конкретном примере. В таблице представлены данные по уловам голубого краба (Callinectes sapidus) в течение 12 недель. Во втором столбце представлены уловы за каждую неделю. Третий столбец содержит данные о числе промысловых усилий, приложенных за неделю. В данном случае под промысловым усилием понимается одно прохождение траулера вдоль залива, где проводился отлов. В четвертом столбце расположены данные об уловах в расчете на одно промысловое усилие. Наконец, в пятом столбце приведены данные о т.н. аккумулированном или суммарном улове. Каждая ячейка этого столбца получается путем сложения значения улова в предыдущую неделю (столбец 2) со значением аккумулированного улова в предыдущую неделю.
Как поступают с этими данными далее? На обычном двумерном графике по оси абсцисс откладывают аккумулированные, или суммарные уловы, а по оси ординат – уловы в расчете на одно промысловое усилие. Получают ряд точек, которые приблизительно выстраиваются вдоль ниспадающей прямой. Далее либо на глаз, либо с помощью более точных методов регрессионного анализа собственно и проводят эту прямую. Точка, в которой регрессионная прямая пересечет ось абсцисс и будет соответствовать общей численности популяции.
2.3. Методы оценки плотности популяций
Как мы уже выяснили, плотность популяции – это число особей в расчете на единицу пространства или объема.
Метод квадратов. Суть его заключается в том, что при помощи рамки определенного размера и формы как бы «вырезается» часть пространства, на/в котором обитают особи изучаемой популяции. Именно таким подходом определяется название метода. Однако данный подход настолько широко и давно используется в экологии, что название «квадрат» сегодня превратилось в своего рода сленг. Квадратом называют практически любое соответствующее орудие для учета организмов – от всевозможных рамок округлой и прямоугольной формы, до батометров, которые доносят на поверхность определенный объем воды, и планктонных сетей, которые также облавливают определенный объем воды. Метод требует соблюдения только двух простых условий: 1) точное знание облавливаемой площади или объема; 2) организмы должны быть относительно неподвижны в момент их подсчета.
До того, как непосредственно применить метод квадратов, необходимо решить два важных вопроса: 1) Квадрат какого размера нужно использовать? 2) Какая форма «квадрата» лучше всего подойдет для проведения исследований? Дать ответ на эти, казалось бы, простые вопросы, не так уж легко. Самый простой способ – изучить литературу по своему вопросу и использовать квадраты того же типа, что используют коллеги. Так, например, при учете плотности размещения деревьев обычно используют квадратные рамки со сторонами 10х10 м; в гидробиологии для учета плотности популяций бентосных организмов используют рамки площадью 0,25 м2 либо дночерпатели с площадью захвата 1/40 м2. Проблема при таком подходе заключается в том, что наши коллеги, увы, также не застрахованы от неправильных походов.
Для нахождения оптимальных параметров квадратов необходимо решить, что же вы, собственно, имеете в виду под этими «оптимальными параметрами». «Оптимальность» может определяться двумя различными факторами: 1) со статистической точки зрения лучшим будет тот квадрат, применение которого дает максимальную статистическую точность. Это означает что при некоторых форме и размерах квадрата исследователь получает наименьшую стандартную ошибку средней величины плотности организмов, либо наиболее узкие доверительные интервалы для этой средней; 2) с точки зрения экологии лучшим будет тот квадрат, использование которого наиболее адекватно отражает свойства изучаемой популяции.
Давайте рассмотрим, какие трудности возникают при выборе формы «квадрата». Здесь исследователи сталкиваются с проблемой т.н. «краевого эффекта». Суть его в том, что при использовании «квадратов» одинаковой площади захвата, но разных по форме, некоторые особи изучаемой совокупности могут недоучитываться, либо наоборот – исследователь может включить в подсчет «лишние» особи. Смысл сказанного представлен на рисунке (рисунок). В принципе, многие исследователи заметили, что длинные прямоугольные «квадраты» лучше, чем круглые или квадратные. Это объясняется гетерогенностью среды обитания. Очевидно, что даже в пределах одного и того же биотопа условия существования в разных его частях будут различаться. В соответствии с распределением этих условий обитания обычно распределяются и живые организмы. Прямоугольные «квадраты» позволяют, таким образом, лучше учесть эту гетерогенность. В итоге исследователь получает более точный со статистической точки зрения результат. Вот простой пример. Были произведены учеты плотности растения Prunella vulgaris при помощи квадратов площадью 1 м2, которые, однако, различались по форме. Были использованы квадраты со сторонами 1х1 м и 4х0,25 м. В таблице представлены полученные результаты:
| Средняя плотность | Дисперсия | Ст. ошибка | 95%-ный доверительный интервал | |
| 1х1 м | 565,3 | 5,94 | ±12,65 | |
| 4х0,25 м | 333,3 | 4,56 | ±9,71 |
Очевидно, что в данной ситуации гораздо более эффективными оказались квадраты прямоугольной формы.
В общем, в каждой конкретной ситуации исследователь сам решает, какой квадрат ему использовать, исходя из предварительных наблюдений, а также имеющихся в его распоряжении времени и материальных средств. Существует также ряд статистических методов, позволяющих определить оптимальные параметры квадратов. Описание этих методов можно найти в специальных в руководствах. Чрезвычайно важным является также вопрос о количестве квадратов, в которых производится подсчет организмов. Способы определения оптимального объема выборки можно найти практически в любой книге по биометрии.
Еще одним из наиболее общих методов оценки плотности популяций является метод линейных трансект. Суть его заключается в том, что в пределах исследуемой территории А выделяется полоса определенной ширины (2W). Исследователь (z) движется вдоль средней линии этой полосы и учитывает все попавшиеся ему объекты (х). Понятно, что объекты эти должны быть 1) достаточно крупными, чтобы быть замеченными. Кроме того, они 2) должны обладать невысокой подвижностью, чтобы не быть учтенными более одного раза. Фактически, такой подход аналогичен применению квадратов, только квадрат здесь имеет очень большие размеры и обладает прямоугольной формой. Движение вдоль трансекты можно выполнять пешком, на автомобиле, лодке, вертолете или самолете. В гидробиологии трансекта часто представляет собой не полосу, а просто линию, направленную, например, перпендикулярно берегу. В таком случае исследователь через определенные промежутки вдоль этой линии (=разреза или полуразреза) отбирает необходимые пробы.
Часто интересующие исследователя объекты обнаружить не так-то просто. Например, практически невозможно увидеть большинство птиц, скрывающихся в зарослях.В данном случае применяют весенние учеты по песням самцов. Т.е. исследователь отмечает, сколько раз слышит песню из разных мест на протяжении определенного маршрута. Это достаточно точный способ, поскольку для птиц характерно территориальное поведение и вероятность повторного учета того же самца крайне мала. Более того, при таких учетах обычно допускается, что количество самок в популяции равно количеству самцов. Поэтому получаемые результаты умножают на два.
Далеко не всегда исследователям так легко определить числе
| <== предыдущая лекция | | | следующая лекция ==> |
| Особенности поведения | | |
Дата добавления: 2021-01-11; просмотров: 1676;
Поиск по сайту
Узнать еще
- B) в угле Интинского месторождения и продуктах его сжигания.
- II. Лексико-грамматические разряды местоимений. Их общая характеристика
- IV. Переход местоимений в другие части речи
- VI.2. Классификация месторождений нефти и газа
- VII.2. Закономерности изменения свойств нефтей и газов на месторождениях
- XIII.4. Прямые методы поисков месторождений нефти и газа.
- А) Философия в системе культуры.
- Автодорожный травматизм занимает третье место в мире среди причин смертности населения.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
