Геометрическая оптика 9 глава
Модификация субстрата путем добавления к нему предварительно облученных фотосенсибилизаторов возможна в реакциях типа IV. При облучении растворов фотосенсибилизаторов в присутствии кислорода в них образуются относительно стабильные продукты фотоокисления, имеющие время жизни от нескольких секунд до нескольких дней. Эти продукты фотоокисления при столкновениях с молекулами субстрата реагируют с ними, давая биологически активные конечные продукты.
Фотохимические реакции, инициируемые светом в тканях человека и животных, могут иметь как положительные, так и отрицательные последствия. Раздел медицины, посвященный лечебному применению или патологическим последствиям действия оптического излучения, называется фото медициной.
Известны терапевтические эффекты оптического излучения, осуществляющиеся за счет поглощения света молекулами, содержащимися в биологических тканях. Так, УФ-В-облучение кожи используется для лечения кожного заболевания — псориаза. Фототерапия без использования экзогенных фотосенсибилизаторов применяется для лечения желтухи новорожденных. Желтуха новорожденных возникает из-за накопления в крови аномально высоких концентраций продукта распада гемоглобина — билирубина. Это связано с тем, что в первые дни жизни у некоторых детей наблюдается недостаток фермента глюкуронилтрансферазы, превращающей плохо растворимый в воде билирубин в его водорастворимое производное. Гидрофобный билирубин склонен накапливаться в клетках мозга, что может приводить к необратимым изменениям в центральной нервной системе. Билирубин имеет максимум поглощения в синей области спектра. Под действием синего света билирубин легко фотоизомеризуется непосредственно в кровеносных сосудах, образуя водорастворимые продукты, легко выводящиеся из организма. Таким образом устраняются патологические последствия гипербилирубинемии.
Существует ряд терапевтических методов, называемых фотохимиотерапией, в которых используется совместное воздействие экзогенных фотосенсибилизаторов и света. Наиболее распространены два вида фотохимиотерапии: ПУВА-терапия кожных заболеваний и фотодинамическая терапия злокачественных новообразований.
В ПУВА-терапии5 в качестве фотосенсибилизаторов используются псоралены в сочетании с УФ-А-облучением кожи. Она эффективна при псориазе, витилиго, кожной Т-клеточной лимфоме и других дерматозах. При ПУВА-терапии в коже человека протекают все IV типа фотосенсибилизированных реакций, наиболее важны из них реакции типов III и IV.
Фотодинамическая терапия (ФДТ) используется для удаления опухолей, доступных для облучения светом. ФДТ основана на применении локализующихся в опухолях фотосенсибилизаторов, повышающих чувствительность тканей при их последующем облучении видимым светом. Часто в качестве фотосенсибилизаторов при ФДТ используют производные гематопорфирина, поглощающие в красной области спектра. Ведущую роль при ФДТ играют реакции типа П. Разрушение опухолей при ФДТ основано на трех эффектах: 1) прямое фотохимическое уничтожение клеток опухоли; 2) повреждение кровеносных сосудов опухоли, приводящее к ишемии и гибели опухоли; 3) ФДТ способно вызывать воспалительную реакцию, мобилизующую противоопухолевую иммунную защиту тканей организма.

1 Ультрафиолетовое излучение с длинами волн менее 200 нм используется редко. Оно поглощается кислородом воздуха, поэтому его действие можно изучать только в специальных условиях, например, в вакууме, из-за этого ультрафиолет с X < 200 нм называют вакуумным.
2 Избирательность действия отличает биологические эффекты света от эффектов ионизирующего излучения. Поглощение квантов рентгеновского или у-излучения осуществляется не молекулами, а атомами и не зависит от того, в состав каких молекул эти атомы входят (см. § 26.4). Поэтому поглощение ионизирующего излучения происходит в основном теми элементами, которых в организме больше. Так как наш организм на 80% состоит из воды, то радиохимические процессы приводят преимуще
ственно к появлению радикалов воды, которые в дальнейшем повреждают белки, нуклеиновые кислоты, липиды и др.
3 Здесь интенсивность выражена в числе фотонов, падающих на 1 м2
за 1 с.
4 Фотолиз — химическая модификация вещества под действием света.
5 Русское название этой терапии возникло путем транслитерации английской аббревиатуры PUVA, произошедшей от Psoralens + UVA-radia-tion.
§ 24.10. Биофизические основы зрительной рецепции
В этом параграфе описаны процессы, протекающие от момента поглощения кванта света молекулой зрительного пигмента до возникновения рецепторного потенциала на клеточной мембране палочки.
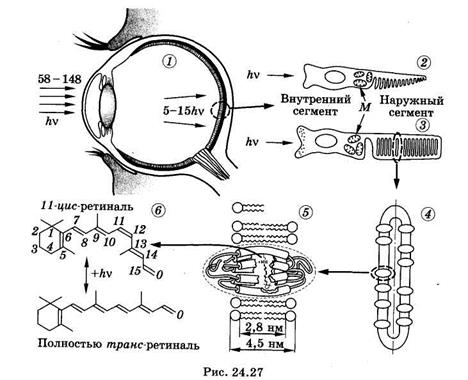
Сначала рассмотрим, как устроен зрительный рецептор. На рйс. 24.27 показаны: 1 — разрез глаза; 2 — колбочка; 3 — палочка (М — скопление митохондрий); 4 — диск наружного сегмента палочки; 5 — фрагмент мембраны диска со встроенной в нее молекулой родопсина; 6 — хромофорная группировка родопсина — ретиналь в 11-цис- и полностью-трамс-конформации. Свет, попавший в глаз (см. § 21.4), фокусируется оптической системой на слой светочувствительных клеток сетчатки — палочек и колбочек. Палочки (их около 125 млн) располагаются по всей поверхности полусферической сетчатки и отвечают за черно-белое, или сумеречное, зрение. Колбочек значительно меньше (примерно 6,5 млн). Они, главным образом, сконцентрированы в центральной части сетчатки, на прямой, проходящей через центры роговицы и хрусталика, и отвечают за цветовое зрение при достаточно ярком освещении.
Строение сетчатки человека и позвоночных животных на первый взгляд может показаться парадоксальным.
Светочувствительные клетки находятся в заднем слое сетчатки. Прежде чем свет попадет на них, он должен пройти через несколько слоев нервных клеток. Мало того, сами палочки и колбочки ориентированы к свету своим внутренним сегментом, не содержащим зрительного пигмента (см. рис. 24.27). Однако такая организация не снижает существенно чувствительности глаза к свету, так как нервные клетки и внутренние сегменты самих светочувствительных клеток прозрачны для видимого света.
Самые ранние исследования зрения касались порога чувствительности глаза. Абсолютный порог чувствительности зрения человека (т. е. минимальная еще обнаруживаемая энергия светового импульса при отсутствии светового фона и в условиях полной световой адаптации) равен 2,1 • 10-17 — 5,7 • 10-17 Дж на поверхности роговицы глаза, что соответствует 58—148 квантам сине-зеленого света. Однако из них только 5—15 квантов поглощаются собственно молекулами зрительного пигмента в палочках сетчатки, а остальные теряются. Около 4% света отражается от роговицы, около 50% поглощается оптическими средами глаза, при этом около 80—85% света, дошедшего до сетчатки, проходит сквозь нее и поглощается в клетках пигментного эпителия. Благодаря пигментному эпителию, подстилающему сетчатку, резко уменьшается количество отраженного и рассеянного от задней стенки глаза света, и тем самым улучшается острота зрения. Так как в сетчатке содержится 125 млн палочек, все кванты при пороговой интенсивности (а их всего 5—15) поглощаются разными палочками.
Рассмотрим строение и функции палочек (см. рис. 24.27). Зрительный пигмент палочек родопсин сосредоточен в наружных сегментах палочек, где он встроен в зрительные диски. Диски представляют собой замкнутые бимолекулярные липидные мембраны, напоминающие собой расплющенные воздушные шары, уложенные в стопку. Наружный сегмент соединен с внутренним сегментом тонкой соединительной ножкой. Во внутреннем сегменте рядом с ножкой сосредоточено большое количество митохондрий, в нем же располагается ядро клетки. В конце внутреннего сегмента, повернутого к свету, находится синаптический контакт с нервным волокном.
Молекулы родопсина пронизывают липидный бислой мембран зрительных дисков. Для нормальной фоторецепции очень важно, чтобы молекулы пигмента могли совершать быструю латеральную диффузию и находились в очень «мягком» окружении в мембране, так как молекулы пигмента после поглощения фотона претерпевают значительные конформационные перестройки. Низкая вязкость липидного слоя обусловлена очень высоким (до 50%) содержанием полиненасыщенной докозагексеновой жирной кислоты (22 : 6 — она содержит 22 атома углерода и 6 ненасыщенных двойных связей). Однако по этой же причине в липидах мембран могут легко активироваться процессы пероксидного окисления, что, по-видимому, лежит в основе ряда заболеваний глаз.
Зрительный пигмент родопсин — сложный белок. Он состоит из гликопротеидной части — опсина и хромофорной группы — ре-тинальдегида, или просто ретиналя. Ретиналь может иметь несколько пространственных изомеров, например 9-цис-ретиналь, 11-цис-ретиналь, полностью-транс-ретиналь и др. Вся совокупность фотохимических превращений родопсина зиждется на двух фундаментальных явлениях. Во-первых, ретиналь под действием света способен переходить в различные цис-транс-изомеры, наиболее важные из которых (11-цис- и полностью-транс-ретиналь) изображены на рис. 24.27. Во-вторых, только одна из этих изомерных форм, а именно 11-цис-ретиналь, структурно соответствует центру связывания ретиналя на опсине и образует с ним прочный комплекс. Другие изомеры таким соответствием не обладают и комплекс хромофор-белок непрочен.
Единственной фотохимической реакцией, которая приводит к появлению ощущения света, в зрительном рецепторе является фотоизомеризация (см. §24.9) 11-цис-ретиналя в полностью-транс-конформацию. После образования транс-формы пространственное соответствие хромофора и опсина нарушается, а это приводит к целой серии конформационных перестроек в молекуле белка, которые сопровождаются изменениями в спектре поглощения зрительного пигмента. Перестройки родопсина завершаются его распадом на опсин и полностью-яузамс-ретиналь. Далее фермент изомераза переводит полностью-транс-ретиналь в 11-цис-ретиналь, после чего происходит его присоединение к опсину и регенерация родопсина.
 Фотопревращения родопсина приводят к электрическому ответу рецепторной клетки. За этим процессом можно проследить путем регистрации электроретинограмм (экстраклеточного отведения потенциалов от сетчатки). Сразу после освещения палочки короткой вспышкой света наблюдается продолжающийся примерно 1 мс ранний рецепторный потенциал (РРП), амплитуда которого растет с увеличением интенсивности вспышки, но не превышает 5 мВ (рис. 24.28). Затем через 1 мс развивается поздний рецепторный потенциал (ПРП). Обращают на себя внимание несколько особенностей рецепторных потенциалов. Во-первых, в отличие от всех других известных клеток на цитоплазматической мембране наружных сегментов палочек потенциал имеет знак (+) внутри и знак (-) снаружи. Под действием света происходит развитие некоторого подобия потенциала действия нервных клеток, но с противоположным знаком. Во-вторых, индуцированный светом сигнал состоит из двух фаз: РРП и ПРП, природа которых совершенно различна. По-видимому, РРП связан с перемещением молекул родопсина во время конформационных перестроек, вызванных освещением. На молекуле этого белка имеются фиксированные заряды, положение которых относительно бислоя липидов после поглощения квантов света меняется, что и является причиной РРП. Следует подчеркнуть, что в возникновении РРП не участвуют никакие процессы изменения ионной проницаемости мембран. После РРП начинаются процессы совершенно иной природы, в которых решающую роль играет движение ионов через клеточную мембрану.
Фотопревращения родопсина приводят к электрическому ответу рецепторной клетки. За этим процессом можно проследить путем регистрации электроретинограмм (экстраклеточного отведения потенциалов от сетчатки). Сразу после освещения палочки короткой вспышкой света наблюдается продолжающийся примерно 1 мс ранний рецепторный потенциал (РРП), амплитуда которого растет с увеличением интенсивности вспышки, но не превышает 5 мВ (рис. 24.28). Затем через 1 мс развивается поздний рецепторный потенциал (ПРП). Обращают на себя внимание несколько особенностей рецепторных потенциалов. Во-первых, в отличие от всех других известных клеток на цитоплазматической мембране наружных сегментов палочек потенциал имеет знак (+) внутри и знак (-) снаружи. Под действием света происходит развитие некоторого подобия потенциала действия нервных клеток, но с противоположным знаком. Во-вторых, индуцированный светом сигнал состоит из двух фаз: РРП и ПРП, природа которых совершенно различна. По-видимому, РРП связан с перемещением молекул родопсина во время конформационных перестроек, вызванных освещением. На молекуле этого белка имеются фиксированные заряды, положение которых относительно бислоя липидов после поглощения квантов света меняется, что и является причиной РРП. Следует подчеркнуть, что в возникновении РРП не участвуют никакие процессы изменения ионной проницаемости мембран. После РРП начинаются процессы совершенно иной природы, в которых решающую роль играет движение ионов через клеточную мембрану.
Особенностью наружных сегментов палочек сетчатки является то, что в покое (в темноте) их цитоплазматическая мембрана имеет высокую проницаемость для ионов натрия, тогда как в покое мембраны нервных и мышечных клеток имеют высокую проницаемость для ионов калия. Асимметрия распределения ионов в палочке по сравнению с внешней средой такая же, как и для других клеток: внутри мало натрия, но много калия. Ведущую роль в открывании натриевых каналов цитоплазматических мембран палочек играет циклический гуанозинмонофосфат, на свету его концентрация падает. Поэтому после поглощения света родопсином натриевые каналы проницаемости закрываются и главным потенциал-образующим ионом становится калий. В результате развивается поздний рецепторный потенциал со знаком (-) внутри клетки.
При слабом свете максимум чувствительности глаза человека расположен в области около 500 нм, что соответствует максимуму поглощения родопсина, содержащегося в палочках. На ярком свету максимум чувствительности смещается к 550 нм, что соответствует максимуму спектра поглощения пигмента в наиболее  распространенном типе колбочек. Пигмент колбочек содержит тот же самый 11-транс-ретиналь, как и родопсин, но белковая часть пигмента отличается, поэтому пигменты колбочек носят название иодопсинов.
распространенном типе колбочек. Пигмент колбочек содержит тот же самый 11-транс-ретиналь, как и родопсин, но белковая часть пигмента отличается, поэтому пигменты колбочек носят название иодопсинов.
При измерении спектров поглощения отдельных колбочек оказалось, что каждая колбочка содержит только один вид иодопсина, а типов колбочек всего три. Иодопсины человека имеют максимумы поглощения при 445, 535 и 570 нм. Поглощение света иодопсинами приводит к появлению РРП в колбочках. Для того чтобы измерить спектр действия РРП, регистрируют вызываемые короткими вспышками яркого монохроматического света амплитуды РРП (рис. 24.29, а). На основании этих измерений строят спектры действия РРП (рис. 24.29, б), совпадающие по форме со спектрами поглощения зрительных пигментов. В сетчатке карпа таким способом обнаружено три типа колбочек с максимумами спектров действия при 462, 529 и 611 нм.
При некоторых генетических заболеваниях нарушается синтез одного из белков иодопсинов, в результате чего не образуется соответствующий пигмент цветного зрения и человек утрачивает способность различать цвета. Эта болезнь называется дальтонизмом.
Роговица и хрусталик помимо того, что они ответственны за формирование изображения на сетчатке, выполняют также роль граничных светофильтров, пропускающих излучение с длинами волн более 300 и 400 нм, соответственно. В обеих структурах при очень больших дозах УФ-облучения происходят фотохимические повреждения, выражающиеся в помутнении и воспалительных реакциях. Главная причина помутнения хрусталика (катаракты) — его фотоповреждение. В присутствии сенсибилизаторов фотоповреждения глаз резко усиливаются. Так, при ПУВА-терапии заболеваний кожи (см. § 24.9) часть сенсибилизатора, естественно, проникает в структуры глаза. УФ-А-облучение сенсибилизированных животных приводило к морфологически выраженным повреждениям роговицы, вещества внутренней камеры глаза, ирисовой диафрагмы и хрусталика. В основе фотосенсибилизированного псораленами повреждения хрусталика глаза лежат окислительные реакции с белками хрусталика человека — кристаллинами. Максимум спектра действия сенсибилизированного псораленами фотоповреждения глаз расположен при 320—340 нм, хвост спектра доходит до 380 нм. Больные, получающие ПУВА-терапию, во время УФ-А-облучения обязательно надевают светозащитные очки, чтобы избежать повреждения глаз. Им также в течение нескольких часов после приема таблеток псораленов нельзя находиться на прямом солнечном свету, содержащем большое количество УФ-А-излучения. Фотоповреждения хрусталика необратимы, так как поврежденные молекулы из него никогда не выводятся.
До сетчатки УФ-излучениё не доходит, поэтому псоралены и другие фотосенсибилизаторы, поглощающие при l < 400 нм, не эффективны. Однако в клетках сетчатки осуществляются фотоповреждения под действием света, поглощенного хромофорной группой зрительных пигментов ретиналем. Ретиналь, входящий в состав зрительных пигментов, расходует энергию электронного возбуждения в процессах цис-транс-изомеризации. В других фотохимических реакциях связанный с опсином 11-цис-ретиналь, по-видимому, практически не участвует. Но свободный полностью-транс-ретиналь является на свету эффективным генератором синглетного кислорода, т. е. способен фотосенсибилизировать реакции типа II (см. § 24.9). Квантовый выход триплетных возбужденных состояний свободного полностью-транс-ретиналя высок; он составляет 0,4—0,7. Максимумы спектров поглощения различных изомеров свободного ретиналя располагаются при 360—380 нм, длинноволновый хвост захватывает фиолетовую область видимого света. Поэтому фиолетовый свет вызывает генерацию триплетных возбужденных состояний ретиналя, что в свою очередь приводит к значительному окислительному повреждению мембран дисков рецепторных клеток. Субстратом окисления являются главным образом остатки полиненасыщенной докозагексеновой жирной кислоты, содержание которой в фосфолипидах мембран рецепторных дисков очень велико. Для защиты мембран дисков от фотосенсибилизированного окисления в них содержится витамин Е (a-токоферол) в высоких концентрациях. Защитный эффект этого витамина обусловлен тем, что он, с одной стороны, способен переводить синглетный кислород в основное состояние, с другой стороны, он является эффективным ингибитором свободно-радикальных реакций цепного окисления ненасыщенных липидов. Фотоповреждение мембран, сенсибилизированное ретиналем, — очень важный негативный эффект, лежащий в основе повреждения сетчатки глаз под действием видимого света. Для защиты глаз можно рекомендовать два способа: применять антиоксиданты, а также пользоваться светозащитными очками, не пропускающими фиолетовый и синий свет. Ношение синих очков (пропускающих синий свет) очень вредно для глаз, так как эти очки, ослабляя интегральный световой поток, стимулируют расширение зрачков. На сетчатку в результате попадает большее количество фиолетового света, и фотосенсибилизируемое свободным ретиналем повреждение усиливается.
Глава 25
Магнитный резонанс
В этой главе, как и в предыдущей, рассматриваются явления, связанные с излучением и поглощением энергии атомами и молекулами.
Магнитный резонанс — избирательное поглощение электромагнитных волн веществом, помещенным в магнитное поле.
§ 25.1. Расщепление энергетических уровней атомов в магнитном поле
В § 13.1, 13.2 было показано, что на контур с током, помещенный в магнитное поле, действует момент силы. При устойчивом равновесии контура его магнитный момент совпадает с направлением вектора магнитной индукции. Такое положение занимает контур с током, предоставленный самому себе. Существенно иначе ориентируются в магнитном поле магнитные моменты частиц. Рассмотрим этот вопрос с позиции квантовой механики.
В § 23.6 отмечалось, что проекция момента импульса электрона на некоторое направление принимает дискретные значения. Чтобы обнаружить эти проекции, необходимо каким-то образом выделить направление Z. Один из наиболее распространенных способов — задание магнитного поля, в этом случае определяют проекцию орбитального момента импульса [см. (23.26)], проекцию спина (23.27), проекцию полного момента импульса электрона [см. (23.30)] и проекцию момента импульса атома LAz [см. (23.37)] на направление вектора магнитной индукции В.
Связь между моментом импульса и магнитным моментом (13.30) и (13.31) позволяет использовать перечисленные формулы для нахождения дискретных проекций соответствующего магнитного момента на направление вектора В. Таким образом, в отличие от классических представлений, магнитные моменты частиц ориентируются относительно магнитного поля под некоторыми определенными углами.
Для атома, например, из (23.37) получаем следующие значения проекций магнитного момента ртг на направление вектора магнитной индукции: 
где 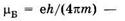 магнетон Бора (см. § 13.1), т — масса электрона, mj — магнитное квантовое число, g — множитель Ланде (g-фактор) (см. § 13.4), для заданного уровня энергии атома он зависит от квантовых чисел L, J, S. Знак «-» в (25.1) обусловлен отрицательным зарядом электрона.
магнетон Бора (см. § 13.1), т — масса электрона, mj — магнитное квантовое число, g — множитель Ланде (g-фактор) (см. § 13.4), для заданного уровня энергии атома он зависит от квантовых чисел L, J, S. Знак «-» в (25.1) обусловлен отрицательным зарядом электрона.
Энергия атома в магнитном поле с учетом того, что в отсутствие поля энергия атома равна Ео, определяется формулой 

Так как магнитное квантовое число mj [см. (23.37)] может принимать 2J + 1 значений от +J до —J, то из (25.2) следует, что каждый энергетический уровень при помещении атома в магнитное поле расщепляется на 2J +1 подуровней. Это схематически показано на рис. 25.1 для J = 1/2. Разность энергий между соседними
подуровнями равна

Расщепление энергетических уровней приводит и к расщеплению спектральных линий атомов, помещенных в магнитное поле. Это явление называют эффектом Зеемана.

Запишем выражение (25.2) для двух подуровней E1 и Е2, образованных при наложении магнитного поля:
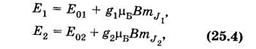
где Е01 и Е02 — энергетические уровни атома в отсутствие магнитного поля. Используя (23.31) и (25.4), получаем выражение для излучаемых атомом частот:
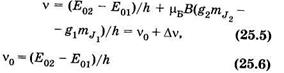
|
где
— — частота спектральной линии в отсутствие магнитного поля;
 — расщепление спектральной линии в магнитном поле. Из (25.7) видно, что Av зависит от магнитного квантового числа, множителя Ланде и магнитной индукции поля. Если g1 = g2 = g, то
— расщепление спектральной линии в магнитном поле. Из (25.7) видно, что Av зависит от магнитного квантового числа, множителя Ланде и магнитной индукции поля. Если g1 = g2 = g, то
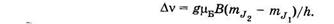
Согласно правилам отбора для магнитного квантового числа, имеем
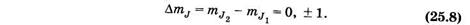
Это соответствует трем возможным частотам: n0 + gmBB/h, n0, n0 - gmBB/h, т. е. в магнитном поле спектральная линия расщепляется и превращается в триплет (рис. 25.2). Такое расщепление называется нормальным или простым эффектом Зеемана. Он наблюдается в сильных магнитных полях или при g1 = g2.
В слабых магнитных полях при g1 ¹ g2 существует аномальный эффект Зеемана, и расщепление спектральных линий значительно более сложное.
§ 25.2. Электронный парамагнитный резонанс и его медико-биологические применения
У атома, помещенного в магнитное поле, спонтанные переходы между подуровнями одного и того же уровня маловероятны. Однако такие переходы осуществляются индуцированно под влиянием внешнего электромагнитного поля. Необходимым условием является совпадение частоты электромагнитного поля с частотой фотона, соответствующего разности энергий между расщепленными подуровнями. При этом можно наблюдать поглощение энергии электромагнитного поля, которое называют магнитным резонансом.
В зависимости от типа частиц — носителей магнитного момента — различают электронный парамагнитный резонанс (ЭПР) и ядерный магнитный резонанс (ЯМР).
ЭПР происходит в веществах, содержащих парамагнитные частицы: молекулы, атомы, ионы, радикалы, обладающие магнитным моментом, обусловленным электронами. Возникающее при этом явление Зеемана объясняют расщеплением электронных уровней (отсюда название резонанса — «электронный»). Наиболее распространен ЭПР на частицах с чисто спиновым магнитным моментом (в зарубежной литературе такую разновидность ЭПР иногда называют электронным спиновым резонансом).
ЭПР был открыт Е. П. Завойским в 1944 г. В первых опытах наблюдалось резонансное поглощение в солях ионов группы железа. Завойскому удалось изучить ряд закономерностей этого явления.
Из выражений (23.31) и (25.3) получаем следующее условие резонансного поглощения энергии: 

Магнитный резонанс наблюдается, если на частицу одновременно действуют постоянное поле индукции Врез и электромагнитное поле с частотой v. Из условия (25.9) понятно, что обнаружить резонансное поглощение можно двумя путями: либо при неизменной частоте плавно изменять магнитную индукцию, либо при неизменной магнитной индукции плавно изменять частоту. Технически более удобным оказывается первый вариант.
На рис. 25.3 показаны расщепление энергетического уровня электрона (а) и изменение мощности Р электромагнитной волны, прошедшей образец, в зависимости от индукции магнитного поля (б). При выполнении условия (25.9) возникает ЭПР.
Форма и интенсивность спектральных линий, наблюдаемых в ЭПР, определяются взаимодействием магнитных моментов электронов, в частности спиновых, друг с другом, с решеткой твердого тела и т. п. Выясним, как эти факторы влияют на характер спектров.
Предположим, что условие (25.9) выполняется. Для поглощения энергии необходимо, чтобы у атомов вещества была большая населенность нижних подуровней, чем верхних. В противном случае, будет преобладать индуцированное излучение энергии.
При электронном парамагнитном резонансе наряду с поглощением энергии и увеличением населенности верхних подуровней происходит и обратный процесс — безызлучательные переходы на нижние подуровни, энергия частицы передается решетке.
Процесс передачи энергии частиц решетке называют спин-решеточной релаксацией, он характеризуется временем т. По соотношению Гейзенберга (23.11) это приводит к уширению уровня.
Таким образом, резонансное поглощение вызывается не точно при одном значении В, а в некотором интервале DВ (рис. 25.4). Вместо бесконечно узкой линии поглощения будет линия конечной ширины: чем меньше время спин-решеточной релаксации, тем больше ширина линии (t1 < t2, соответственно кривые 1 и 2 на рис. 25.4).
Уширение линий ЭПР зависит также от взаимодействия спинов электронов (спин-спиновое взаимодействие) и от других взаимодействий парамагнитных частиц. Разные типы взаимодействия влияют не только на ширину линии поглощения, но и на ее форму.
Поглощенная при ЭПР энергия, т. е. интегральная (суммарная) интенсивность линии, при определенных условиях пропорциональна числу парамагнитных частиц. Отсюда следует, что по измеренной интегральной интенсивности можно судить о концентрации этих частиц.
Важными параметрами, характеризующими синглетную (одиночную) линию поглощения, являются npeз, Врез, g (положение точки резонанса), соответствующие условию (25.9). При постоянной частоте v значение Врез зависит от g-фактора. В простейшем случае g-фактор позволяет определить характер магнетизма системы (спиновый или орбитальный). Если же электрон связан с атомом, входящим в состав твердой кристаллической решетки или какой-либо молекулярной системы, то на него будут влиять сильные внутренние поля. Измеряя g-фактор, можно получить информацию о полях и внутримолекулярных связях.
Однако если бы при исследовании получалась только синглетная линия поглощения, то многие приложения магнитных резонансных методов были бы невозможны. Большинство приложений, в том числе и медико-биологических, базируется на анализе группы линий. Наличие в спектре ЭПР группы близких линий условно называют расщеплением. Имеется два характерных типа расщепления для спектра ЭПР.
Первое — электронное расщепление — возникает в тех случаях, когда молекула или атом обладают не одним, а несколькими электронами, вызывающими ЭПР. Второе — сверхтонкое расщепление — наблюдается при взаимодействии электронов с магнитным моментом ядра.
 Современная методика измерения ЭПР основывается на определении изменения какого-либо параметра системы, происходящего при поглощении электромагнитной энергии.
Современная методика измерения ЭПР основывается на определении изменения какого-либо параметра системы, происходящего при поглощении электромагнитной энергии.
Прибор, используемый для этой цели называют ЭПР-спектрометром. Он состоит из следующих основных частей (рис. 25.5): 1 — электромагнит, создающий сильное однородное магнитное поле, индукция которого может плавно изменяться; 2 — генератор СВЧ-излучения электромагнитного поля; 3 — специальная
«поглощающая ячейка», которая концентрирует падающее СВЧ-излучение на образце и позволяет обнаружить поглощение энергии образцом (объемный резонатор); 4 — электронная схема, обеспечивающая наблюдение или запись спектров ЭПР; 5 — образец; 6 — осциллограф.
Дата добавления: 2020-11-18; просмотров: 260;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
