Характеристика антител (иммуноглобулинов)
Несмотря на многообразные неспецифические механизмы иммунной защиты организма, он не мог бы быть надежно защищен от антигенов, если бы не существовали механизмы специфической иммунной защиты, поскольку антигены эволюционировали параллельно с эволюцией иммунной системы и выработали ряд механизмов, позволяющих некоторым из них
Ø избегать прямого контакта с фагоцитами (резко изменив пространственную структуру своих поверхностных рецепторов)
Ø полностью избежать активации системы комплемента по альтернативному пути
Ø либо после активации системы комплемента по альтернативному пути и образования на поверхности патогена опсонизирующего фагоцитоз фактора С3b все равно не фагоцитироваться фагоцитами.
Эволюционным предкам человека необходимо было выработать механизмы, которые служили бы для защиты от каждого конкретного микроорганизма, как бы много таких микроорганизмов ни было. Формирование таких механизмов было обеспечено благодаря развитию специфических средств иммунной защиты. Такие специфические средства иммунной защиты могут обеспечиваться
Ø либо антителами, которые продуцируются плазматическими клетками (потомками В-лимфоцитов) и строго специфически взаимодействуют с определенными антигенами, являющимися, как правило, внеклеточными паразитами,
Ø либо Т-лимфоцитами, которые обеспечивают уничтожение внутриклеточных паразитов и собственных генетически измененных клеток организма.
Антитела (иммуноглобулины) представляют собой белковые молекулы, которые строго специфически взаимодействуют с определенными антигенами и
Ø либо непосредственно их обезвреживают,
Ø либо облегчают последующий фагоцитоз антигенов фагоцитами,
Ø либо активируют систему комплемента по классическому пути, в результате чего образуется мембранноатакующий комплекс, вызывающий повреждение антигена.
Несмотря на большое разнообразие антител, все они имеют общий план строения. В частности, в основе антител лежит Y-образная молекула, состоящая из четырех цепей: двух легких и двух тяжелых. Тяжелые цепи более длинные и занимают центральное положение в молекуле, а легкие цепи относительно короткие и прикрепляются снаружи к верхним участкам (N-концевым участкам) тяжелых цепей, тогда как нижние участки тяжелых цепей (С-концевые участки) образуют "хвост" молекулы. Тяжелые цепи соединены между собой и с легкими цепями с помощью дисульфидных связей (S-S-связей). Антигенраспознающими участками антитела являются верхние его плечи, образованные верхними (N-концевыми) частями легкой и тяжелой цепей.
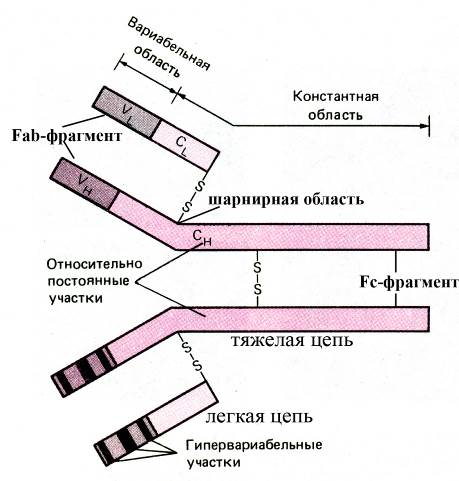
Рис. Структура иммуноглобулина G
В антигенраспознающих участках тяжелой и легкой цепей молекулы антитела, в свою очередь, различают:
Ø 3последовательно расположенные гипервариабельные области (CDR-области, от англ. Complementarity Determining Regions), эти области определяют специфичность антител, их аминокислотный состав сильно варьирует у различных антител. Именно эти области тяжелой и легкой цепей специфически (по принципу стереохимического соответствия) взаимодействуют с определенными антигенами. Причем антиген оказывается расположенным внутри плеча определенного антитела, т.е. окружен, с одной стороны гипервариабельными областями легкой, а с другой – тяжелой цепей. В гипервариабельных областях легких цепей обнаруживается большое количество аминокислотных остатков глицина, обуславливающих гибкость этих участков полипептидной молекулы. Существует предположение, согласно которому специфичность к антигену обеспечивается гипервариабельными участками тяжелой цепи, а "тонкая настройка" достигается благодаря хорошей конформационной гибкости легкой цепи
Ø и соединяющие эти гипервариабельные области промежуточные пептидные последовательности (аминокислотный состав которых мало отличается у разных антител), называемые каркасными областями (framework-областями). Каркасные области обеспечивают не только связь между гипервариабельными областями внутри цепи, но взаимодействие с каркасными участками вариабельного домена другой цепи (между вариабельными доменами легкой и тяжелой цепей).
Хвост молекулы антитела, образованный нижними (С-концевыми) частями двух тяжелых цепей, не обладает специфичностью (в молекулах антител, относящихся к одному классу, концевые части тяжелых цепей имеют сходное строение) и ответственен за связывание с рецепторами собственных клеток организма. Следовательно, у каждой молекулы антитела имеется как минимум:
ü два антигенраспознающих и соответственно антигенсвязывающих участка, соответствующих плечам молекулы и называемых Fab-фрагментами (от англ. fragment antigen binding)
ü и один неспецифичный фрагмент, обеспечивающий взаимодействие антитела с рецепторами собственных клеток организма и соответствующий хвосту молекулы антитела, образованному дистальными частями тяжелых цепей (т.н. Fc-фрагмент, получивший название от англ. fragment crystallizable).
В тяжелых цепях молекулы антител у места перехода плечей в хвост (т.е. в области "шейки" молекулы) содержится большое количество аминокислотных остатков пролина, что обеспечивает конформационную гибкость молекулы и необходимо для лучшего взаимодействия с антигенными детерминантами, находящимися на поверхности клеток. Область тяжелых цепей, соответствующая переходу плеча тяжелой цепи в хвост и обладающая высокой конформационной гибкостью, называется шарнирной областью антитела.
В зависимости от особенностей строения (аминокислотной последовательности) константных областей тяжелых цепей, молекулы иммуноглобулинов (Ig) классифицируются на 5 классов (или изотипов):
ü А(в их состав входят тяжелые цепи a-типа),
ü G (в их состав входят тяжелые цепи g-типа),
ü M(в их состав входят тяжелые цепи m-типа),
ü D (в их состав входят тяжелые цепи d-типа),
ü E (в их состав входят тяжелые цепи e-типа).
На основании особенностей строенияконстантных областей легких цепей иммуноглобулинов выделяют 2 разновидности (2 изотипа) легких цепей: c и l, причем в состав определенной молекулы антитела всегда входят идентичные легкие цепи (либо обе c-цепи, либо обе l-цепи). Таким образом, в пределах каждого класса иммуноглобулинов в зависимости от того, какие изотипы легких цепей входят в состав молекулы антитела, можно выделить два типа антител (например, иммуноглобулины класса G представлены двумя типами молекул: Gc и Gl, а класса М – Мc и Мl и т.д.).
Тяжелые и легкие цепи антител имеют сложную пространственную структуру. В частности, они состоят из последовательно расположенных глобулярных доменов, соединенных между собой линейными участками (состоящими приблизительно из 20 аминокислотных остатков). Каждый глобулярный домен имеет вид петли, которая объединяет в своем составе до 60 аминокислот и образуется в результате замыкания дисульфидных связей между определенными аминокислотными остатками цистеина внутри какой-то из цепей антитела.
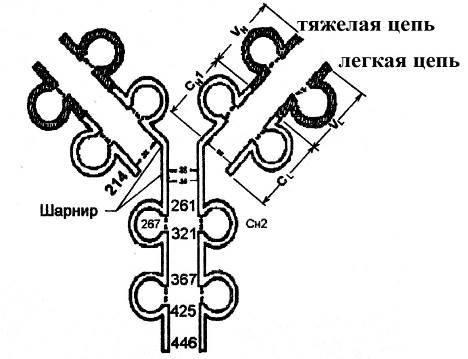
Рис. Принцип доменной организации молекулы иммуноглобулина (на примере иммуноглобулина G). Каждый домен включает приблизительно 100-110 аминокислотных остатков; причем около 60 аминокислотных остатков домена оказываются заключенными в петлю дисульфидной (S-S-связью); около 20 аминокислотных остатков домена, которые не входят в состав петли, служат для соединения с другими доменами. Цифры обозначают последовательность аминокислотных остатков в полипептидных цепях. VL и CL – вариабельный и константный домены легкой цепи. VH – вариабельный домен тяжелой цепи, CH1 CH2 CH3 – константные домены тяжелой цепи.
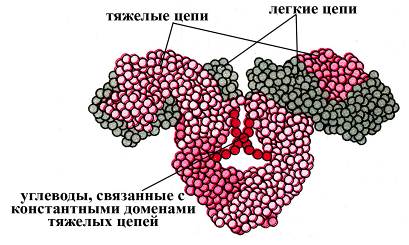
Рис. Компьютерная модель иммноглоблина G
Внутри доменов пептидные фрагменты, входящие в состав домена, образуют компактно уложенную антипараллельную b-складчатую структуру, стабилизированную водородными связями (вторичная структура белка). Образованию b-складчатой структуры внутри домена способствуют аминокислотные остатки глицина. Таким образом, части тяжелых и легких цепей антител внутри доменов формируют b-складчатые структуры (вторичная структура белка), которые, в свою очередь, укладываются с образованием петлеобразных доменов (третичная структура белка). Благодаря слоистой b-складчатой структуре внутри домена три гипервариабельные области в каждой цепочке антитела оказываются максимально приближенными друг к другу.
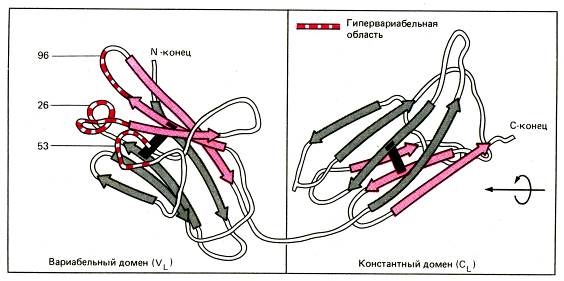
Рис. Структура глобулярных доменов(вариабельного и константного)легкой цепи (по данным рентгеноструктурного анализа белков Бенс-Джонса). Одна поверхность каждого домена состоит из 4-х цепей (серые стрелки), образующих антипараллельную b-складчатую структуру, стабилизированную межцепочечными водородными связями (между группами СО и NH на протяжении всего пептидного остова). Другая поверхность каждого домена образована тремя цепями (розовые стрелки). Полипептидные цепи, образующие две поверхности домена связаны друг с другом межцепочечной дисульфидной связью (обозначена самой темной полосой). Описанная структура характерна для всех иммуноглобулиновых доменов. Особый интерес представляет расположение гипервариабельных областей в трех отдельных петлях вариабельного домена (гипервариабельные области обозначены красно-белыми полосатыми линиями, цифрами обозначены некоторые аминокислотные остатки в гипервариабельных областях). Эти гипервариабельные области, хотя и находятся на большом отдалении друг от друга в первичной структуре легкой цепи, но при образовании пространственной структуры они оказываются расположенными в непосредственной близости друг к другу, принимая участие в формировании антигенсвязывающего центра иммуноглобулина.
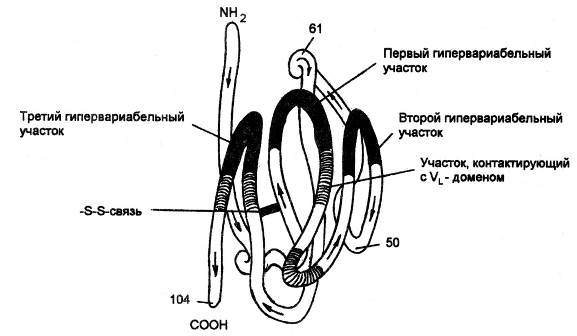
Рис. Пространственное расположение гипервариабельных участков внутри вариабельного домена тяжелой цепи иммуноглобулина G человека. Конформационная особенность вариабельного домена состоит в том, что все 3 гипервариабельных участка в результате формирования третичной структуры полипептидной цепи оказываются в непосредственной близости друг от друга (черные участки рисунка). Каркасные (инвариантные) участки обеспечивают взаимодействие с каркасными участками вариабельного домена легкой цепи. В результате взаимодействия вариабельного домена легкой и тяжелой цепей и формируется антигенсвязывающий центр иммуноглобулина.
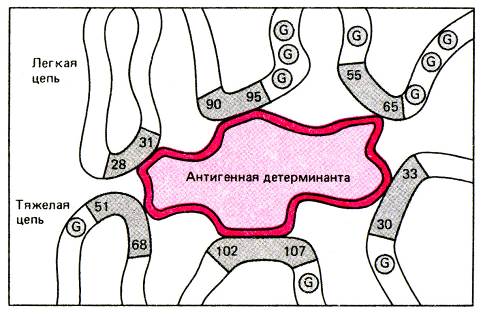
Рис. Упрощенное двумерное изображение атигенсвязывающго центра антитела. Антигенсвязывающий центр антитела представляет собой полость, окруженную пептидными петлями гипервариабельных участков тяжелой и легкой цепей (на рисунке пронумерованы аминокислотные остатки гипервариабельных областей цепей).
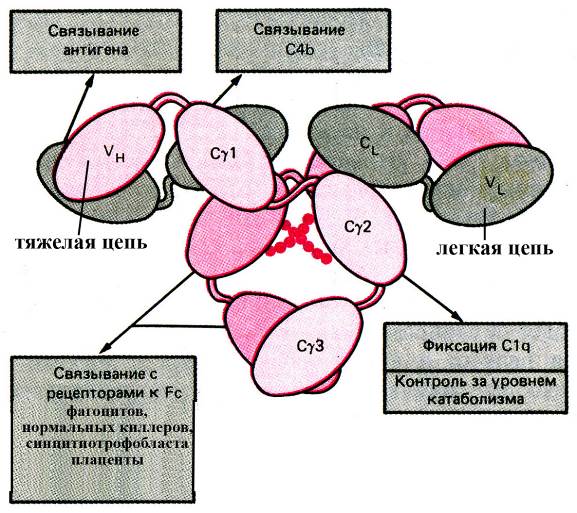
Рис. Функциональное значение различных доменов иммуноглобулина G (схема). Домены легкой цепи обозначены буквами VL (вариабельный домен) и CL (константный домен); домены тяжелой цепи g-типа обозначены VH (вариабельный домен тяжелой цепи) и Сg1 Сg2 и Сg3 (константные домены тяжелой цепи)
Как уже было сказано выше, на основании особенностей строения константных областей тяжелых цепей молекул иммуноглобулинов, выделяют 5 их классов, каждый из которых характеризуется определенными особенностями организации Fc-фрагментов, обуславливающими то, с рецепторами каких эффекторных клеток организма будет взаимодействовать такой иммуноглобулин и некоторые другие его функциональные особенности. Преобладающим классом иммуноглобулинов во внутренних жидкостях организма (и преимущественно в тканевой жидкости) являются антитела класса G, которые в больших количествах продуцируются при вторичном иммунном ответе и обеспечивают защиту организма от бактерий, вирусов и токсинов. В частности, комплексы "IgG-антиген"
· усиливают фагоцитоз посредством опсонизации (т.е. комплексы "IgG-антиген" Fc-фрагментами IgG взаимодействуют с рецепторами мембраны нейтрофилов и макрофагов, повышая эффективность фагоцитоза антигенов),
· стимулируют внеклеточное уничтожение антигенов путем активации естественных киллеров (IgG, связанные с антигенами, своими Fc-фрагментами способны взаимодействовать не только с фагоцитами, но и с естественными киллерами, повреждающими мембрану антигена)
· обладают способностью взаимодействовать с компонентом С1 системы комплемента, активируя ее по классическому пути, что сопровождается появлением
ü медиаторов воспаления, обладающих хемотаксическим действием и привлекающих фагоциты и лимфоциты,
ü опсонизирующего фагоцитоз фактора С3b
ü и в конечном итоге образованием мембраноатакующих комплексов, разрушающих патогены.
Интенсивность синтеза IgG во многом зависит от проникновения антигенов в организм. IgG является единственным антителом, способным проникать через плацентарный барьер, поскольку на поверхности клеток трофобласта плаценты расположены рецепторы, связывающие Fc-фрагменты молекул материнских IgG. При этом связанные с рецепторами трофобласта молекулы IgG поглощаются путем опосредованного рецепторами эндоцитоза, после чего транспортируются в клетке трофобласта в составе окаймленных пузырьков, выводятся из клеток трофобласта, проходят через базальную его мембрану в соединительную ткань и капилляры плода. Переход IgG через плаценту обеспечивает передачу пассивного иммунитета от матери к плоду. Кроме того, в связи с присутствием IgG в молоке, он принимает участие в пассивной специфической иммунной защите ребенка в период грудного вскармливания.
Иммуноглобулины класса А представляют собой основной класс иммуноглобулинов в секретах экзокринных желез (молочных, слезных, слюнных, потовых желез, желез слизистой оболочки пищеварительной трубки и бокаловидных клеток дыхательной и мочеполовой трубок). IgА выделяется на поверхность слизистых оболочек, где и взаимодействует с антигенами. Следовательно, IgА участвует в защитной функции организма, укрепляя барьер в слизистой оболочке пищеварительного тракта, дыхательной и мочеполовой трубок против инфекций. Молекула иммуноглобулина А, содержащаяся в составе секрета каких-то желез представляет собой димер, стабилизированный дополнительной J-цепью. Причем димерные молекулы IgА образуются в самой плазматической клетке. После чего димерные молекулы IgА взаимодействуют с определенными полипептидными рецепторами на базальной поверхности секреторной клетки. IgА-рецепторы в комплексе с димером IgА путем эндоцитоза проникают внутрь секреторной клетки и наряду с опосредованием фагоцитоза секреторной клеткой, обеспечивают защиту IgА от расщепления под действием протеолитических ферментов синтезируемых клеткой секретов. После секреции IgА через апикальную поверхность секреторной клетки на поверхность слизистой оболочки рецептор для IgА частично расщепляется и его часть, оставшаяся связанной с димером IgА после такого расщепления, носит название секреторного компонента. IgА играет важную роль в защите слизистых оболочек от инфекций, которая обеспечивается его способностью препятствовать проникновению связанных с IgА микроорганизмов через эпителиальный пласт слизистой оболочки в ткани. В плазме крови молекулы IgА имеют преимущественно мономерное строение.
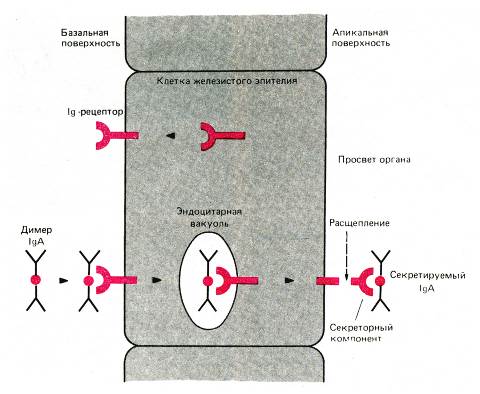
Рис. Механизм секреции иммуноглобулина А на поверхность слизистой оболочки. Эпителиальные клетки слизистой синтезируют иммуноглобулиновый рецептор (Ig-рецептор), который встраивается в мембрану базальной поверхности клетки. Димер Ig А связывается с этим рецептором, путем эндоцитоза, проникает внутрь клетки, транспортируется к апикальной ее поверхности, через мембрану которой путем экзоцитоза выводится на поверхность слизистой оболочки. При расщеплении рецептора на поверхности слизистой оболочки высвобождается Ig А, который все еще связан с частью рецептора, получившей название секреторного компонента. Транспорт Ig G через плаценту возможно происходит подобным образом с помощью рецепторов для Ig G, расположенных на поверхности клеток трофобласта.
Иммуноглобулины класса Е содержатся в сыворотке крови человека, как правило, в небольших концентрациях, обладают способностью своими Fc-фрагментами взаимодействовать с рецепторами тучных клеток и базофилов. После присоединения к поверхностной мембране тучных клеток и базофилов комплексов "IgЕ-антиген" или взаимодействия фиксированных на поверхности мембраны этих клеток молекул IgЕ с определенными антигенами происходит дегрануляция тучных клеток и базофилов, сопровождающаяся выделением в окружающую тканевую жидкость гистамина (расширяет артериолы, повышает проницаемость капилляров, в больших дозах суживает венулы) и гепарина (ингибирует образование нерастворимого фибрина и его отложение на внутренней поверхности сосудистой стенки, тем самым повышая проницаемость капилляров). Выброс этих медиаторов воспаления базофилами и тучными клетками обуславливает многие проявления воспалительных и аллергических реакций, привлекая в очаг воспаления неспецифические и специфические защитные агенты, что способствует уничтожению различных паразитов (в связи с этим уровень IgЕ в крови возрастает не только при аллергиях, но и при паразитарных инвазиях). Таким образом, основная физиологическая функция IgЕ, очевидно, состоит в защите внешних слизистых оболочек организма путем локальной активации факторов крови и эффекторных клеток благодаря индукции острой воспалительной реакции. Инфекционные агенты, способные прорвать линию обороны, созданную IgА, будут связываться со специфическими IgЕ на поверхности тучных клеток, в результате чего тучные клетки начнут дегранулировать и высвобождать свои секреты (вазоактивные амины и хемотаксические факторы) в окружающую ткань, что, в свою очередь, вызовет приток циркулирующих в крови IgG, компонентов комплемента, нейтрофилов и эозинофилов в инфицированный очаг. В этих условиях способность эозинофилов повреждать гельминтов, нагруженных IgG, и усиленная продукция IgЕ в ответ на проникновение этих паразитов в организм будут обеспечивать эффективную защиту.
Иммуноглобулины класса М представляют собой пентамерные молекулы (т.е. состоят из пяти Y-образных субъединиц, расположенных радиально и объединенных в единую молекулу с помощью единственной J-цепи, которая через посредство дисульфидных связей взаимодействует с тяжелыми цепямимономеров). При этом Fc-фрагменты каждого мономера обращены к центру молекулы и друг к другу, а Fab-фрагменты – кнаружи. J-цепь и инициирует сборку пентамерной молекулы IgМ. В связи с пентамерным строением своих молекул IgМ имеют самую большую среди всех классов антител молекулярную массу (950 кД).
 Иммуноглобулины М представляют собой первый класс антител, продуцируемых развивающимися В-лимфоцитами при первичном попадании антигена в организм и содержащихся в наибольших количествах в периферической крови (т.е. IgМ образует первую линию обороны при бактериемии). IgМ благодаря большому размеру своей молекулы в комплексе с антигеном способен в единичном количестве активировать компонент С1 системы комплемента, запуская процесс активации этой системы по классическому пути, тогда как для активации компонента С1 комплексом "IgG-антиген" необходимо присоединение к его молекуле 5 комплексов " IgG-антиген".
Иммуноглобулины М представляют собой первый класс антител, продуцируемых развивающимися В-лимфоцитами при первичном попадании антигена в организм и содержащихся в наибольших количествах в периферической крови (т.е. IgМ образует первую линию обороны при бактериемии). IgМ благодаря большому размеру своей молекулы в комплексе с антигеном способен в единичном количестве активировать компонент С1 системы комплемента, запуская процесс активации этой системы по классическому пути, тогда как для активации компонента С1 комплексом "IgG-антиген" необходимо присоединение к его молекуле 5 комплексов " IgG-антиген".
Кроме активации системы комплемента, IgМ оказывает опсонизирующее действие при фагоцитозе. Более того, в связи с пентамерным строением IgМ способен вызывать агглютинацию и обусловленный этим лизис антигенов. Теоретически молекула IgМ может связать 10 антигенов, но, как правило, эффективно взаимодействует только с 5-ю, что обусловлено определенными стерическими ограничениями, возникающими из-за недостаточной гибкости молекулы. Мономерные молекулы IgМ представлены на поверхности В-лимфоцитов, образуя рецепторы для взаимодействия с антигеном.
Рис. Структура иммуноглобулина М
Иммуноглобулины класса D присутствуют в сыворотке крови в ничтожно малых количествах, но зато преимущественно связаны с мембраной лимфоцитов и, очевидно, выступают в роли рецепторов лимфоцитов, позволяющих им взаимодействовать между собой, благодаря чему обеспечивается контроль за активацией и супрессией лимфоцитов.
Дата добавления: 2016-07-18; просмотров: 5121;
Поиск по сайту
Узнать еще
- Cравнительная характеристика усилителей на БТ
- I. Загальна характеристика
- I.3. Антитела системы АВ0
- II. Монозы и их характеристика
- Nemathelmintes. Общая характеристика типа. Nematoda. Характеристика класса. Медицинское значение. Био- и геогельминты.
- Plahelmintes (Плоские черви).Общая характеристика типа. Морфология, систематика, основные представители, значение.
- Protozoa. Общая характеристика подцарства. Классификация. Представители. Медицинское значение.
- V. Механическая характеристика

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
