Репродуктивные органы цветковых растений
Покрытосеменные, или цветковые растения являются наиболее крупным отделом растительного царства, насчитывающим более 350 семейств, 13 тысяч родов и до 240 тысяч видов. В настоящее время покрытосеменные распространены по всему Земному шару – от тропиков до приполярных пустынь. Автотрофные цветковые растения – важнейший компонент биосферы, от которого зависят биосферные процессы обмена веществ и трансформации энергии, газовый состав атмосферы, климат, водный режим суши, характер процессов почвообразования. На суше цветковые часто доминируют в растительном покрове, только в немногих сообществах и типах растительности они уступают господство по общей массе другим группам растений – в хвойных лесах голосеменным, в некоторых типах тундр и на сфагновых болотах – мохообразным.
Покрытосеменные определяют самую возможность существования большинства наземных животных. Они формируют среду обитания для разнообразных консументов – потребителей первичного органического вещества. Различные группы гетеротрофов связаны с ними трофическими, консортивными связями. В процессе эволюции многие группы гетеротрофов смогли возникнуть только тогда, когда Землю завоевали покрытосеменные и образуемые ими сообщества. Для многих членистоногих и позвоночных характерна сопряженная с цветковыми эволюция. Человек как биологический вид тоже возник и эволюционировал после появления покрытосеменных, отличающихся высоким уровнем продуктивности и биохимическим разнообразием.
Покрытосеменные растения отличаются необычайным полиморфизмом, эволюционной пластичностью и могут существовать в разнообразных условиях обитания. Это единственная группа среди высших растений, представители которой сумели вторично освоить морскую среду, где стали доминировать на огромных акваториях.
Разнообразие размеров и жизненных форм цветковых растений, их приспособлений к конкретным условиям обитания велико. Представитель семейства рясковых – ряска малая(Lemna minor) не превышает 3-5 мм в длину и похожа на свободноплавающие слоевищные растения – мхи или водоросли (рис. 13.1).

Рис. 13.1. Ряска малая (Lemna minor) – миниатюрное цветковое растение.
С другой стороны, гигантские эвкалипты(виды родаEucalyptusиз семейства миртовыеMyrtaceae) вырастают высотой до 100 м, длина побегов тропических лиан – ротанговых пальм измеряется сотнями метров. Среди покрытосеменных преобладают автотрофы, но встречаются полупаразиты, паразиты и сапротрофы, полностью утратившие хлорофилл и способность к фотосинтезу. Среди покрытосеменных имеются также эпифиты и эпифиллы (эпифиты, живущие на листьях других растений), деревья, кустарники, кустарнички, полукустарники и травы (многолетники и однолетники), плотоядные растения с их разнообразными приспособлениями для улавливания и переваривания мелких животных(рис. 13.2).

Рис. 13.2. Насекомоядное цветковое растение росянка круглолистная (Drosera rotundifolia).
Отмечены разнообразные приспособления видов цветковых растений к опылению строго определенными агентами, к распространению плодов и семян, к перенесению неблагоприятных климатических периодов. Эти особенности дают цветковым растениям возможность полностью реализовать свой эволюционно-адаптационный потенциал. Ни одна другая группа растений не смогла выработать такого разнообразия приспособлений к определенным факторам среды.
Существенным признаком покрытосеменных считают цветок – отсюда одно из названий отдела – Anthophyta (antos– цветок,phyton– растение). Происхождение цветка как особого органа – это одна из ключевых проблем морфологии и филогении покрытосеменных. Дискуссии по этому вопросу, начавшиеся еще вXVIIIв., продолжаются и в настоящее время.
Типичный цветок определяют как видоизмененный побег, обеспечивающий функции спороношения, развития гаметофитов, половое размножение, образование семян в плоде. В истории ботаники отмечено, что в поисках архетипа цветка за первичный примитивный цветок различные ученые принимали разные структуры.
Сторонники эвантовойтеории считают первичным крупный обоеполый цветок, опыляемый насекомыми, с большим и часто неопределенным числом свободных частей, и тогда цветки однополые, олигомерные, голые рассматриваются как результат более поздних этапов редукционной эволюции.
Сторонники псевдантовойтеории полагают, что эволюционные трансформации выражались в постепенном усложнении структуры цветка от однополых, анемофильных, с небольшим числом частей к обоеполым, полимерным, энтомофильным, с хорошо развитым околоцветником.
Следует учитывать, что в процессе эволюционного возникновения покрытосеменные сначала приобрели комплекс важнейших общих признаков (в первую очередь – уникальные особенности жизненного цикла), а уже потом выработали разные цветки. При этом в каждой эволюционной ветви покрытосеменных были осуществлены различные морфогенетические решения.
Существенными частями цветка являются андроцей, состоящий из одной, нескольких или многих тычинок (микроспорофиллов) и гинецей, который состоит из одного, нескольких или многих плодолистиков, или карпелл (мегаспорофиллов).
Плодолистики (карпеллы) могут быть свободными, и тогда каждый из них, срастаясь краями, образует пестик (один или несколько пестиков в цветке – по числу плодолистиков). При срастании нескольких плодолистиков формируется ценокарпный гинецей, пестик в нем всегда один.
Важнейшая часть пестика – завязь. Наличие завязи (замкнутой полости), в которой развиваются семязачатки (в отличие от голосеменных, у которых семязачатки расположены на поверхности открытых мегаспорофиллов), обычно считают важнейшим признаком покрытосеменных, откуда и другое название отдела – Angiospermae(от греческогоangeion – сосуд, вместилище,sperma– семя). Полагают, что завязь обеспечивает лучшую защиту семяпочек (семязачатков). В полости завязи семязачатки оказываются в условиях влажной камеры, оптимальных для их развития. Благодаря образованию завязи расширяются и обогащаются возможности распространения растений: из цветка формируется плод, при диссеминации расселяются не только отдельные семена, но и целые плоды, у которых возникают разнообразные приспособления к распространению широким кругом агентов.
У цветковых растений и некоторых голосеменных спермин (мужские неподвижные половые клетки) доставляются к яйцеклетке с помощью особой клетки – пыльцевой трубки, которая обеспечивает высокую точность попадания спермия в зародышевый мешок (то есть к женскому гаметофиту с яйцеклеткой). Семязачатки у покрытосеменных растений скрыты в завязи пестика, поэтому необходимо приспособление для улавливания пыльцы. Таким аппаратом у покрытосеменных служит рыльце на верхушке столбика пестика (стилодия). Наличие рыльца – важная особенность покрытосеменных растений.
Архегониальные растения (папоротникообразные, голосеменные) характеризуются тенденцией к возникновению разноспоровости, которая сопровождается редукцией полового поколения в жизненном цикле. Покрытосеменные, у которых редуцированы гаметангии, представляют собой завершающий этап на этом пути. Предельное сокращение процессов микро- и мегаспорогенеза и ускорение развития гаметофитов, связанное с их редукцией, были, вероятно, важными эволюционными факторами. Женский гаметофит цветковых растений представлен зародышевым мешком, состоящим всего из 7 клеток, а мужской гаметофит представлен пыльцевым зерном (пылинкой), в которой только 2 клетки – сифоногенная и спермагенная (дающая 2 спермия).
Уникальной особенностью покрытосеменных является двойное оплодотворение, в котором участвуют 4 клетки: два спермия, яйцеклетка и центральная клетка зародышевого мешка. В ходе этого процесса один из cnepмиев оплодотворяет яйцеклетку, образуя диплоидную зиготу, которая впоследствии развивается в зародыш семени. Второй спермий сливается с диплоидным ядром центральной клетки зародышевого мешка, образуя триплоидную зиготу. Эта зигота впоследствии дает начало триплоидному эндосперму семени. Общий для всех покрытосеменных уникальный процесс двойного оплодотворения указывает на их эволюционное единство и монофилетическое становление.
Одно из следствий двойного оплодотворения – формирование питательной ткани – полиплоидного эндосперма (в отличие от гаплоидного эндосперма голосеменных, представляющего остаток женского гаметофита). Биологическое значение полиплоидного состояния запасающей ткани связано с тем, что оно обеспечивает более благоприятные условия для развития зародыша и поэтому могло иметь важную эволюционную роль.
На тканевом уровне подавляющее большинство покрытосеменных характеризуется наличием в ксилеме сосудов, тогда как почти у всех голосеменных проводящие элементы древесины представлены только трахеидами. Только у немногих цветковых проводящая система ксилемы гомоксильная, бессосудистая, унаследовавшая примитивность от предковых голосеменных форм.
Паренхиматизация цветковых позволила выработать специальные запасающие ткани и травянистые жизненные формы. В отличие от древесных растений, у трав возможно сокращение онтогенеза и ускорение темпов эволюции; травянистые растения способны существовать в таких крайних условиях среды, в которых деревья и кустарники жить не могут.
Остается нерешенной проблема геологического возраста цветковых растений. Считалось, что покрытосеменные растения возникли раньше мелового периода – в триасе или палеозое (255-210 млн. лет назад). Предполагалось, что первоначально небольшая, эта группа существовала в таких условиях, которые не благоприятствовали фоссилизации их остатков, например, в горных районах или в пустынях. Позже, в меловой период (145-63 млн. лет назад), покрытосеменныедали мощную вспышку развития и сформировали свое современное разнообразие.
Этому взгляду противоречат объективные данные палеоботаники. Многочисленные указания на нахождение ископаемых остатков покрытосеменных в домеловых осадочных породах в настоящее время достоверно не доказаны. Имеются косвенные указания на возможность существования покрытосеменных в юре (210-145 млн. лет назад). Например, с этого времени известны некоторые формы насекомых, современные родственники которых настолько тесно связаны с цветковыми растениями, что допустить их независимое от последних существование трудно.
Древнейшие, достоверно определяемые ископаемые остатки цветковых растений в виде фоссилизованных листьев, цветков, плодов, древесины и пыльцевых зерен встречаются лишь с начала мелового периода (135 млн. лет назад). В этих отложениях они очень немногочисленны и фрагментарны. Еще недавно считалось, что к середине мелового периода (120 млн. лет назад) покрытосеменные появились в массе и выступали как «победители в борьбе за существование» с голосеменными и папоротникообразными. Сейчас палеоботаники отодвигают начало расцвета покрытосеменных на более позднее время – конец мелового периода (80-63 млн. лет назад).
Предполагают, что родиной цветковых могли быть тропические или субтропические регионы Земли. Исследователи считают, что это были районы тропической Юго-Восточной и Восточной Азии и Меланезия. В этом районе сосредоточены древние и считающиеся примитивными таксоны цветковых. Вероятно, что отсюда шла миграция древнейших цветковых в другие области Земли. Еще одна точка зрения заключается в признании центром происхождения покрытосеменны[ древнего континента Южного полушария – Гондваны. Считается, что цветковые могли возникнуть и расселиться по этому обширному континенту до начала раскола Гондваны на современные континенты Южного полушария.
К началу XX в. сложились две противостоящие друг другу системы взглядов на происхождение покрытосеменных от более примитивных предковых форм растений. Одна из них, связанная с именами австрийского ботаника Р. Веттштейна и классика немецкой ботаники А. Энглера, была основана на представлении о происхождении цветковых от эфедроподобных или гнетоподобных голосеменных предков. Р.Веттштейном была разработана оригинальная концепция происхождения цветка – так называемая псевдантовая гипотеза (от греческого «псевдантиум» – ложный цветок). Согласно этой гипотезе цветок представляет собой собрание мелких разнополых стробилов, претерпевших в процессе эволюции ряд преобразований: редукцию, сближение и срастание. На основе этой гипотезы наиболее архаичными следует считать семейства с тычиночными и пестичными невзрачными ветроопыляемыми цветками (ивовые, казуариновые).
Другая концепция восходит к построениям И.В.Гёте и А.П.Декандоля, то есть к концу XVIII – началу XIX веков. В эпоху филогенетической систематики это направление ознаменовалось появлением стробилярной (от греческого «стробилюс» – шишка хвойного)гипотезы, которую сформулировали Е.Арбер и Дж. Паркин . Другое название этой гипотезы – эвантиевая (от греческого «эвантиум» – настоящий цветок). Согласно стробилярной гипотезе наиболее вероятными предками покрытосеменных были мезозойские голосеменные растения – беннеттиты. С этих позицийцветок – это метаморфизированный (видоизмененный) укороченный спороносный побег, первоначально напоминавший шишку голосеменных. Мегаспорофиллы, несущие семязачатки, в процессе метаморфоза превратились в плодолистики (карпеллы), а несущие пыльники микроспорофиллы – в тычинки. Эти видоизменения многие исследователи связывают с приспособлением покрытосеменных к насекомоопылению (энтомофилии). Примитивные цветки имели значительные размеры и выпуклое цветоложе, на котором располагались многочисленные микро- и мегаспорофиллы. Микроспорофиллы эволюционировали в тычинки, а мегаспорофиллы – в карпеллы (плодолистики), которые образовали пестики. Цветки приобрели строение, сходное с тем, которое наблюдается у современных магнолиевых, лютиковых, кувшинковых. Поэтому перечисленные семейства обычно располагаются в основании системы покрытосеменных как наиболее древние и архаичные.Дальнейшая эволюция цветка в пределах покрытосеменных имела редукционный характер.
Обе эти гипотезы являются фолиарными (от латинского «фолиум» – лист), так как исходят из представлений, что цветки образовались из листостебельных спороносных побегов. Фолиарным гипотезам противопоставляются различные теломные (от греческого «телома» – конечные побеги) гипотезы (например, теория гонофилла Р. Мелвилла), широко распространенные в 60-70-е годы ХХ в. Согласно этим гипотезам, все части цветка могут быть выведены из теломов – осевых структур, свойственных ископаемым растения – риниевым.
Палеоботаника в последние десятилетия дала много нового материала о древнейших, ранне- и среднемеловых покрытосеменных. Эти данные позволяют сделать несколько заключений. Во-первых, уже самые ранние из известных нам цветковых отличаются большим полиморфизмом. Очень разнообразны пыльцевые зерна по форме, размерам, скульптуре поверхности, характеру поровости, числу и строению апертур. Они обнаруживают сходство с пыльцевыми зернами порядков и семейств современных растений. Листья меловых цветковых также весьма разнообразны и напоминают современные фикусы, протейные, дубы, акации, аралию, подорожник, известны и специализированные щитовидные листья.
Большим полиморфизмом отличаются и органы размножения – цветки и плоды. У меловых цветковых известны как одиночные и относительно крупные цветки, так и мелкие, собранные в разнообразно устроенные соцветия с простым или двойным околоцветником, с апокарпным или ценокарпным гинецеем, с верхней или нижней завязью, с разным числом тычинок и плодолистиков. Наряду со вскрывающимися плодами типа листовок обнаружены и не-вскрывающиеся плоды. Вместе с древесными формами в меловую эпоху существовали многочисленные травянистые покрытосеменные.
Во-вторых, если раньше часто утверждали, будто в меловую эпоху покрытосеменные были представлены современными таксонами, то сейчас большинство исследователей считают, что раннемеловыеAngiospermae принадлежали к вымершим порядкам и семействам. Этот факт важен как доказательство того, покрытосеменные не оставались неизменными с момента зарождения.
В целом сейчас большинство специалистов считают, что предков Angiospermaeнужно искать среди каких-то древних и малоспециализированных голосеменных растений. Не оставлена идея их генетической связи с беннеттитовыми. Допускают, что магнолиевые (как наиболее примитивные цветковые) лишь сохранили наибольшее число архаичных признаков, но сами, как и все прочие таксоны цветковых, произошли от какой-то вымершей, еще более древней гипотетической группы покрытосеменных.
В настоящем учебном пособии принята система цветковых растений, разработанная А.Л.Тахтаджяном в 1987 г. В данной системе древнейшей группой покрытосеменных считается порядок магнолиевых (Magnoliales), от предков которых, вероятно, произошли современные покрытосеменные. В этом смысле приведенная система монофилетична (рис. 13.3).

Рис. 13.3.Система покрытосеменных растений (по А.Л.Тахтаджяну).
Цветок – особый побег покрытосеменных растений, ответственный за формирование спор двух типов (микро- и мегаспор), развитие мужского и женского гаметофитов, образование гамет (яйцеклеток и спермиев), опыление, оплодотворение и формирование семян. Цветок в процессе своего развития превращается в плод с семенами. Во многих отношениях он уникален и столь характерен для этой группы, что покрытосеменные нередко называют цветковыми растениями. Исключительная роль цветка как особой морфологической структуры связана с тем, что в нем полностью совмещены все процессы бесполого и полового размножения. В цветках осуществляются микро- и мегаспорогенез, развитие гаметофитов, микро- и мегагаметогенез, опыление, оплодотворение и формирование зародышей нового поколения спорофитов. Завершается онтогенез цветка образованием плода с семенами. Особенности строения цветка обеспечивают осуществление всех этих функций с наименьшими затратами пластических веществ и энергии.
Морфология цветка
Цветок разнообразен у разных групп растений по деталям строения, окраске и размерам. Известны цветки до 1 мм в диаметре (рясковые – Lemnaceae) и одновременно существуют очень крупные цветки, например у знаменитой раффлезии Арнольда (Rafflesia arnoldii). Крупнейший цветок этого вида растений имеет диаметр более 100 см.
Цветок возникает из конуса нарастания цветочного побега. Листочки околоцветника, тычинки и пестики последовательно образуются в виде бугорков из верхушечной меристемы. Первоначально процессы формирования и развития цветочных структур осуществляются в цветочной почке. Цветочная почка обычно состоит из почечного покрова (перулы), образованного почечными чешуями – видоизмененными листьями, которые плотно окружают молодой цветок (бутон). Иногда покров отсутствует, тогда бутон защищают молодые листья, плотно облегающие отдельные цветки или целые соцветия.
Существуют два типа цветочных почек: собственно цветочные почки (бутоны), которые помимо чешуй несут только репродуктивные части, и смешанные почки, несущие репродуктивные части и зачатки вегетативного побега. Положение частей цветочного покрова в бутоне относительно друг друга называется листосмыканием. Известно довольно большое число типов листосмыкания. Главнейшие из них – створчатое, свернутое и черепитчатое. Тип листосмыкания покровов в бутоне – важный систематический признак. Его легко удается отразить на диаграмме цветка.
По положению цветок бывает верхушечным или боковым, то есть выходит из пазухи видоизмененного или невидоизмененного прицветного листа (прицветника). Часть побега между цветком и прицветником называют цветоножкой. Если цветоножка укорочена или отсутствует, то цветок называют сидячим. Верхняя часть оси цветка, к которой прикрепляются все боковые (латеральные) его части, является цветоложем. Цветоложе может иметь различные размеры и форму: удлиненную, выпуклую, плоскую или вогнутую. Разросшаяся часть цветоложа, расположенная между покровами цветка и гинецеем, называется тором. Части цветка принято делить на стерильные – к ним относятся покровы цветка, или околоцветник, и фертилъные, то есть репродуктивные (андроцей и гинецей).
У некоторых цветков в результате срастания цветоложа, нижних частей покрова и андроцея образуется особая структура, называемая гипантием. Она может быть разнообразной формы, иногда участвует в формировании плода (у шиповника) и характерна для представителей семейства розоцветных и многих видов бобовых.
Части цветка располагаются на цветоложе в виде ряда концентрических кругов (циклический цветок) или спирально (ациклический цветок). Иногда наружные листочки околоцветника расположены кругами, а внутренние по спирали. Такой цветок является гемициклическим (полуциклическим). Эволюционно ациклические цветки архаичнее (древнее) циклических.
В зависимости от количества частей в каждом круге цветок называют: мономерным, или одночленным, если в одном круге одна часть (ива Salix); димерным, или двучленным – с двумя частями в круге (капустныеBrassicaсеае); тримерным, или трехчленным (многие однодольные); тетрамерным, или четырехчленным с четырьмя членами в круге; пентамерным, или пятичленным.
Иногда число кругов и частей (членов) в них увеличивается (особенно у садовых форм). Такой цветок называют махровым. Махровость обычно связана с расщеплением лепестков в онтогенезе цветка, либо с превращением в лепестки тычинок, что повышает декоративные качества сортов культурных роз, пионов, гвоздик. По особенностям симметрии цветки делят на актиноморфные, или правильные цветки (через цветок можно провести несколько плоскостей симметрии) и зигоморфные (цветок делится плоскостью симметрии на правую и левую половины). В процессе приспособления к опылению с помощью насекомых из актиноморфных цветков возникли цветки зигоморфные (цветки многих бобовых). Существует также асимметричные цветки, через которые нельзя провести ни одной плоскости симметрии (цветки валерианы лекарственной Valeriana officinalis).
Околоцветник, или стерильная часть цветка, является его покровом, защищающим более нежные репродуктивные части. Он бывает простым и двойным. Простой околоцветник состоит из относительно сходных листочков и не разделен на чашечку и венчик. Невзрачный простой околоцветник называют чашечковидным, яркоокрашенный – венчиковидным.
Число долей простого околоцветника у разных систематических групп неодинаково. Оно неопределенно у ряда примитивных семейств из подклассов магнолиид и ранункулид. В цветках однодольных число долей околоцветника обычно кратно трем. Иногда околоцветник частично или полностью редуцирован, что, как полагают, связано с приспособлением к ветроопылению. Большое значение при характеристике цветка имеет тип симметрии околоцветника (рис. 13.4).

Рис. 13.4.Симметрия цветка.А – актиноморфный (радиально симметричный), или правильный цветок; Б – зигоморфный (двусторонне симметричный), или неправильный цветок.
Двойной околоцветник дифференцирован на чашечку и венчик, обычно существенно различающиеся по окраске и размерам. Чашечкой называют совокупность чашелистиков. Чашелистики чаще всего зеленые, разнообразные по форме и образуют наружный или несколько наружных кругов околоцветника. Число чашелистиков в цветке варьирует от двух (семейство маковых Papaveraceae) до неопределенного числа (семейство чайныхTheaceae), но у большинства двудольных их чаще четыре или пять. Чашелистики могут быть совершенно свободными (несросшимися); в таких случаях чашечка называется раздельнолистной. Если же они срастаются между собой на большем или меньшем их протяжении, то чашечка называется сростнолистной. В сростнолистной чашечке выделяют трубку и отгиб (зубцы, лопасти) в зависимости от характера и степени срастания чашелистиков. По форме трубки различают трубчатую, колокольчатую и воронковидную чашечки. Чашечка возникла эволюционно в результате концентрации вокруг цветка и видоизменения верхних прицветных листьев. Главная функция чашечки связана с защитой цветка на ранних этапах его формирования. Наружные покровы бутона до его распускания у цветков с двойным околоцветником образованы чашечкой. При распускании цветка и во время цветения чашечка иногда опадает (семейство маковыеPapaveraceae) или отгибается назад. Нередко она способна видоизменяться, приобретая иные функции, связанные с распространением плода и семян. В семействе яснотковых (Lamiaceae) чашечка служит вместилищем для дробного плода, у астровых она превращена в хохолок (паппус), способствующий разносу плодов ветром. У череды на чашечке имеются характерные крючочки, сохраняющиеся при плодах; с их помощью плоды цепляются за шерсть животных. Иногда чашечка приобретает яркую окраску и выполняет роль венчика, который в этом случае нередко оказывается редуцированным (живокостьDelphiniumи аконитAconitumиз семейства лютиковыхRanunculaceae).
Венчик образует внутреннюю часть двойного околоцветника. Эволюционно листочки венчика произошли в результате стерилизации и общей редукции тычинок (пыльников), уплощения их тычиночных нитей и приобретения ими окраски.
По разнообразию размеров, формы и окраски венчик превосходит остальные части цветка и создает его облик. Венчик образован лепестками, число которых может быть неопределенным, но чаще равно четырем, пяти или трем, реже двум либо одному. В отдельных случаях венчик полностью редуцирован, тогда его функции переходят к чашечке. Венчик содействует опылению цветка, привлекая насекомых-опылителей. Эта роль выполняется относительно пассивно благодаря окраске, размерам или характерной форме, но у некоторых специализированных групп цветковых венчик способен активно участвовать в процессе опыления (бобовые, орхидные), способствуя более успешной его реализации. Благодаря яркой окраске лепестков венчик способен отражать часть спектра солнечных лучей, предохраняя репродуктивные части цветка от перегрева. Закрываясь на ночь, венчик, создает камеру, препятствующую охлаждению цветка или повреждению его холодной росой.
Окраска венчика может значительно варьировать иногда у одного и того же вида. В тропиках более обычны растения с красной, оранжевой либо сине-фиолетовой окраской венчика. В странах умеренного климата преобладает желтый цвет. Окраску определяют различные пигменты полифенольной природы (флавоноиды, в том числе антоцианы, придающие красный, синий или фиолетовый цвет, и флавоны – желтый). В данном случае окраска зависит от активной реакции (рН) клеточного сока (красный цвет обусловлен кислой реакцией, фиолетовый – нейтральной, синий – щелочной). Оранжевую, желтую, красную окраску лепестков иногда обусловливают также каротиноиды. Иногда (семейство бурачниковых Boraginaceae) синяя или красная окраска обусловлена пигментами – беталаинами. Обесцвечивание или потеря пигментов называется альбинизмом. Помимо окраски, воспринимаемой человеком, на венчике существуют узоры, различаемые насекомыми, воспринимающими ультрафиолетовую часть спектра. Эти узоры, возможно, дают насекомым дополнительную информацию о размещении нектарников или пыльников в цветке, способствуют их посещению и опылению.
Пластинка лепестка чаще недифференцирована, но иногда четко подразделяется на две части: нижнюю – суженную, получившую название ноготка, и верхнюю, называемую пластинкой, или отгибом (семейства гвоздичные Caryophyllaceae, бобовыеFabaceae)(рис. 13.5).
Обычно все лепестки венчика более или менее одинаковы, но у некоторых специализированных семейств различаются по величине и форме (фиалковые Violaceae, бобовыеFabaceae, бальзаминовыеBalsaminaceae). Встречаются случаи образования особых полых выпячиваний у отдельных лепестков – так называемых шпорцев (иногда они возникают из листочков простого околоцветника), связанных с особенностями опыления. В полости шпорца накапливается нектар, выделяемый его стенками или специальными нектарниками.
Различают раздельно- и сростнолепестные венчики. Древнейшие из ныне живущих цветковых (магнолиевые, лютиковые, кувшинковые, пионовые) имеют раздельнолепестные венчики. Сростнолепестность в процессе морфологической эволюции возникла позднее и независимо в разных филетичеcких линиях покрытосеменных. Сростнолепестный венчик часто имеется у насекомоопыляемых цветковых. В сростнолепестных венчиках различают нижнюю сросшуюся часть, называемуютрубкой, и верхнюю расширенную – отгиб. Место перехода трубки в отгиб называют зевом.
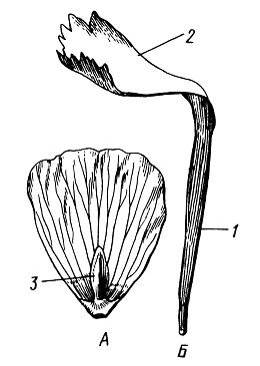
Рис. 13.5.Лепестки.А – сидячий (лютик едкийRanunculus acris); Б – ноготковый (гвоздикаDianthus versicolor): 1 – ноготок, 2 – отгиб, 3 – чешуйка, прикрывающая нектарную ямку.
В зеве иногда располагаются различного рода выросты и придатки – дополнительные приспособления к перекрестному опылению (семейства бурачниковые Baraginaceae, гвоздичныеCaryophyllaceae, горечавковыеGentianaceae). У нарциссов (Narcissus) выросты простого венчиковидного околоцветника очень крупные, ярко окрашенные и образуют как бы второй «венчик», называемый привенчиком, или коронкой. Длина трубки венчика различна и отражает особенности опыления разных видов растений. Увеличение длины трубки, которая у некоторых тропических видов достигает 20-25 см, связана с приспособлением к опылению длиннохоботковыми бабочками и птицами (колибри). Венчики и весь околоцветник разнообразны по форме: трубчатые, язычковые, блюдцевидные, двугубые (рис. 13.6).
Рис. 13.6.Формы сростнолепестных венчиков. А – колесовидный (вербейникLysimachia vulgaris); Б – воронковидный (табакNicotiana tabacum); В – колокольчатый (некоторые однодольные); Г – трубчатый (подсолнечникHelianthus annuus); Д – трубчатый с блюдцевидным отгибом (сирень обыкновеннаяSyringa vulgaris); Е – колпачковый (виноградVitis vinifera); язычковый (одуванчик лекарственныйTaraxacumofficinale); 3 – двугубый (большинство представителей губоцветных, многие норичниковые): 1 – трубка венчика, 2 – отгиб, 3 – зев венчика
Совокупность тычинок одного цветка называют андроцеем. В некоторых цветках андроцей вообще отсутствует – это так называемые пестичные цветки, а в однополых тычиночных цветках имеются только тычинки. Количество тычинок в цветке у разных видов различно – от одной (орхидные Orchidaceae) до нескольких сотен (подсемейство мимозовыеMimosoideae, семейство бобовыеFabaceae). Многочисленные тычинки магнолиевых (Magnoliaceae), имеющих полимерный андроцей, расположены по спирали. У примитивных видов тычинки расположены группами или в несколько кругов. Для большинства цветковых характерно 3, 4, 5, 6 или 10 тычинок (олигомерный андроцей). Обычно они расположены в 1-2 круга. Эволюция шла от полимерного андроцея к олигомерному. Тычинки могут быть свободными либо срастаются различным образом и в разной степени. Например, в тропическом семействе мелиевых все 10 тычинок срастаются своими нитями в трубку (однобратственный андроцей). У зверобоя (Hypericum) тычинки срастаются в пучки, для астровых (Asteraceae) характерно склеивание пыльников. У многих представителей подсемейства мотыльковыхFaboideae (семейство бобовыеFabaceae) срастаются 9 тычинок, а одна остается свободной (двубратственный андроцей) (рис. 13.7).
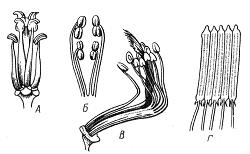
Рис. 13.7.Типы андроцея. А – четырехсильный (капустные, или крестоцветныеBrassicaceae); Б – двусильный (яснотковые, или губоцветныеLamiaceae); В – двубратственный (бобовыеFabaceae); Г – со склеенными в трубку пыльниками (астровые, или сложноцветныеAsteraceae)
Каждая тычинка состоит из суженной нитевидной или лентовидной части – тычиночной нити и расширенной части – пыльника. Пыльник имеет две половинки, соединенные друг с другом связником, являющимся продолжением тычиночной нити. Связник иногда продолжен в надсвязник, заметный в виде небольшого выступа над пыльником. Длина тычиночных нитей у разных растений варьирует. Чаще они более или менее равны по длине околоцветнику, но иногда значительно короче или во много раз его превышают, как, например, у тропического лекарственного растения «почечный чай», или кошачьи усы (Orthosiphon stamineus) из семейства губоцветных (Lamiaceae). На поперечном срезе через тычиночную нить видно, что большая ее часть состоит из паренхимной ткани, а в центре проходит сосудистый пучок.
Каждая половинка пыльника несет два (реже одно или много) гнезда, которые являются микроспорангиями. Гнезда пыльников называют пыльцевыми мешками. В зрелом пыльнике перегородки между гнездами по большей части исчезают. Снаружи пыльники покрыты эпидермой. Под эпидермой располагается слой клеток эндотеция со вторично утолщенными клеточными оболочками, за счет которого при подсыхании пыльника вскрываются гнезда. Глубже залегают 1-3 слоя некрупных тонкостенных клеток. Самый внутренний слой клеток, выстилающий полость пыльцевых мешков, получил название тапетума. Считается, что содержимое клеток этой ткани служит питанием для развивающихся материнских клеток микроспор (микроспороцитов). Гнезда пыльника обычно заполнены материнскими клетками микроспор, микроспорами и зрелой пыльцой. Микроспоры возникают из микроспороцитов в результате мейоза, сами микроспороциты – из немногих клеток археспория (образовательной ткани, функционирующей на ранних стадиях развития гнезд пыльника). Созревший пыльник вскрывается разнообразно: продольными трещинами, дырочками, клапанами. При этом пыльца высыпается. Признаки строения, формы, положения, числа тычинок, а также сам тип андроцея имеют большое значение для систематики цветковых и познания их филогении.
У некоторых видов часть тычинок утратила свою первоначальную функцию, они стали в процессе эволюции стерильными и превратились в стаминодии. Иногда пыльники преобразуются в нектарники – секреторные части цветка, выделяющие нектар. Превратиться в нектарники могут также лепестки, их части, части пестика и даже выросты цветоложа. Нектарники различаются по форме, располагаются обычно в глубине цветка и нередко выделяются своей блестящей поверхностью. В пыльниках осуществляется два важнейших для репродукции процесса: микроспорогенез и микрогаметогенез. В результате из микроспоры формируется зрелый мужской гаметофит (мужскую особь) – пыльцевое зерно. Эволюционно тычинки – это видоизмененные микроспорофиллы ископаемых голосеменных – предков цветковых растений.
Совокупность плодолистиков одного цветка, образующих один или несколько пестиков, называют гинецеем (от греческого «гине» – женщина). Плодолистики, или карпеллы – это структуры, связанные по происхождению со спороносными листьями (мегаспорофиллами) ископаемых предков цветковых растений. В ходе эволюции из плоских и открытых мегаспорофиллов возникли вдоль сложенные (кондупликатные) плодолистики, края которых затем срослись и образовали замкнутый пестик с его наиболее существенной частью – завязью, несущей на внутренней поверхности семязачатки. У цветковых из мегаспорофиллов эволюционно сформировалась уникальная структура, имеющая замкнутую полость, в которой развиваются надежно защищенные от внешних воздействий семязачатки. Нижняя часть пестика – завязь выполняет функцию влажной камеры, предохраняющей семязачатки от высыхания, что сделало покрытосеменные малозависимыми от влажности окружающей среды и явилось одним из факторов освоения ими аридных (за
Дата добавления: 2016-05-30; просмотров: 4852;
Поиск по сайту
Узнать еще
- Адаптация растений и животных к световому режиму.
- Анализаторы как органы ощущений
- Анатомическое строение стебля однодольных и двудольных растений. Строение стебля двудольных и голосеменных древесных растений. Структура древесины
- Ассортимент растений, рекомендуемый для создания санитарно-защитных зон и озеленения города
- Афферентный сигнал. Афферентный нерв. Исполнительные органы. Обратная афферентация ( связь ).
- Б) Адаптации растений к тепловому режиму
- Б) Экологические группы растений по отношению к свету
- Бактериальный фотосинтез и его типы. Отличие бактериального фотосинтеза от фотосинтеза растений.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
