Способы объединения фрагментов ДНК
Создание рекомбинантной ДНК – это процесс объединения in vitro двух или более фрагментов ДНК, выделенных из различных биологических источников. Фрагменты ДНК, содержащие нуклеотидные последовательности структурных или регуляторных участков генов, представляющих интерес для исследователя, получают с использованием эндонуклеаз рестрикции. Эти ферменты, как было рассмотрено в предыдущей главе, могут образовывать фрагменты ДНК как с тупыми, так и с липкими концами. В зависимости от цели, для которой создается новая ДНК, а также от наличия в арсенале исследователя ферментов для модификации ДНК, объединение различных фрагментов ДНК производится тремя основными методами, зависящими от того, какие концы имеют фрагменты сшиваемых ДНК.
Лигирование по одноименным "липким" концам
Этот метод является самым распространенным и популярным. Впервые этим способом гибридная ДНК была получена С. Коэном с сотрудниками в 1973 году. Одноцепочечные концы, получаемые при гидролизе ДНК эндонуклеазами рестрикции, например EcoRI, образуют водородные связи с комплементарными им последовательностями другого конца этого же фрагмента или другого фрагмента ДНК. Таким образом, любые два фрагмента (независимо от их происхождения), образовавшиеся под действием одной и той же эндонуклеазы рестрикции, могут слипаться за счет образования водородных связей между комплементарными основаниями нуклеотидов однонитевых концевых участков (рис. 13).
Однако, образование водородных связей между комплементарными основаниями одноцепочечных участков ДНК не восстанавливает полной целостности двойной спирали, поскольку остаются два разрыва в сахарнофосфатном остове полинуклеотидной цепи. Для восстановления фосфодиэфирных связей в полинуклеотидных цепях ДНК, т.е. для сшивания, или лигирования нитей, используют фермент ДНК-лигазу.
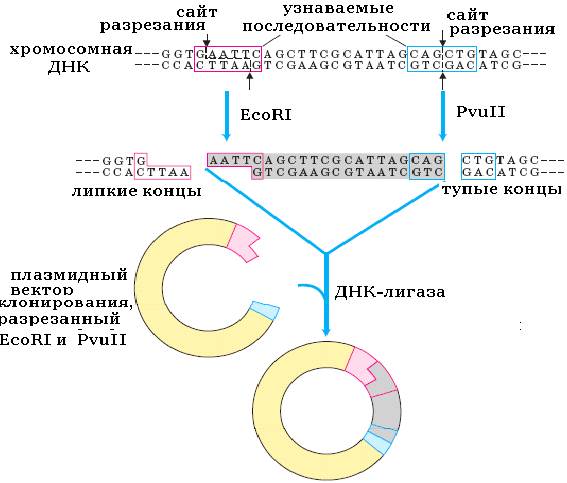
Рис. 13. Схема объединения фрагментов ДНК по липким и тупым концам (рестрикционно-лигазный метод).
Этот фермент в живой клетке выполняет ту же функцию - сшивание фрагментов ДНК, синтезирующихся при репликации.
Лигирование по одноименным "липким" концам с обработкой щелочной фосфатазой
При лигировании двух фрагментов ДНК, один из которых является линеаризованным вектором (см. раздел векторы), а другой – донорной ДНК, содержащей нужный фрагмент ДНК и предварительно обработанной такой же эндонуклеазой как и вектор, целесообразно применить еще один фермент-инструмент: щелочную фосфатазу. Чтобы предупредить комплементарное взаимодействие между липкими концами одного и того же типа рестрицированных фрагментов, например – линеаризованного вектора, проводят обработку фосфатазой, которая отщепляет фосфатные группы на 5’-концах одноцепочечных липких концов. ДНК-лигаза не может сшить концы дефосфорилированной линейной векторной ДНК (рис. 14).
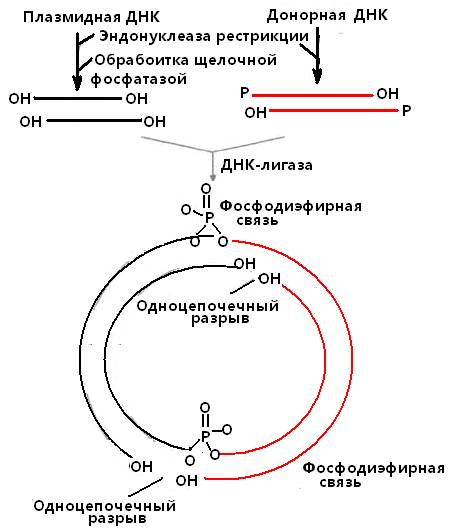
Рис. 14 . Встраивание чужеродной ДНК в плазмидный вектор.
Плазмидную ДНК, обработанную эндонуклеазой рестрикции и щелочной фосфатазой, смешивают с рестрицированной донорной ДНК, содержащей нужный ген, и добавляют ДНК-лигазу. Два из четырех одноцепочечных разрыва при этом устраняются, и конструкция оказывается стабильной благодаря образовавшимся фосфодиэфирным связям
Что касается собственно рекомбинантных молекул ДНК, то хотя в них и имеются два одноцепочечных разрыва, ее фрагменты удерживаются вместе двумя фосфодиэфирными связями, образовавшимися с помощью ДНК-лигазы между дефосфорилированной плазмидной ДНК и рестрицированной донорной ДНК (рис. 14). При репликации этой ДНК после введения ее в клетку-реципиент одноцепочечные разрывы устраняются системой лигирования клетки-хозяина.
Лигирование по "тупым" концам
Тупые концы также можно соединенять с помощью ДНК-лигазы, если и фермент, и тупые концы присутствуют в реакционной смеси в высоких концентрациях (рис. 13) В этом случае реакция лигирования имеет свои особенности и ее эффективность ниже, чем при сшивке по липким концам. Впервые такие эксперименты были выполнены в 1972 году Полем Бергом в Стенфордском университете, США.
Липкие концы можно присоединить к фрагментам ДНК с тупыми концами ферментативным путем. Для этого используют фермент терминальную трансферазу из тимуса теленка, который присоединяет нуклеотиды к 3’-концам цепей ДНК. Если к 3'-концам одного из рекомбинируемых in vitro фрагментов ДНК с помощью концевой дезоксинуклеотидилтрансферазы достроить одноцепочечные олиго (dA)-сегменты определенной длины, а к концам другого фрагмента — олиго (dT)-сегменты примерно такой же длины, то при объединении полученных таким образом фрагментов происходит комплементарное спаривание за счет образования водородных связей между олиго (dА)- и олигo (dT) -последовательностями (рис. 15). Для ковалентного соединения двух фрагментов используется ДНК-лигаза. Эти процедуры составляют основу для второго общего метода получения рекомбинантных молекул ДНК, который получил название коннекторный метод.
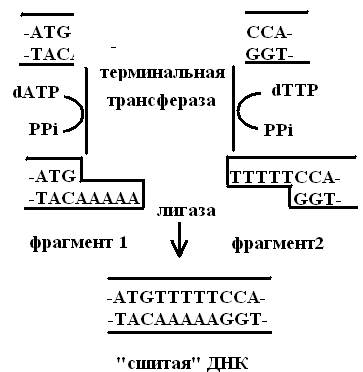
Рис. 15. Создание «липких» концов и сшивка фрагментов ДНК коннекторным методом
Поскольку можно формировать достаточно длинные взаимокомплементарные одноцепочечные концы, гибридные молекулы образуются с высокой эффективностью. В частности, поэтому при клонировании ДНК-копий матричных РНК, которые доступны в ограниченных количествах, обычно используют коннекторный метод. При таком способе соединения между фрагментами встраиваются участки ААААА. Такие дополнительные последовательности ТТТТТ могут влиять на функции соединяемых молекул и поэтому всегда, когда только возможно, для получения рекомбинантных молекул ДНК пользуются липкими концами, образовавшимися в результате действия эндонуклеаз рестрикции.
Дата добавления: 2016-07-18; просмотров: 5574;
Поиск по сайту
Узнать еще
- I. Способы представления переменного синусоидального тока и напряжения.
- MS Word. Выделение текста. Понятие фрагмента текста. Способы форматирования фрагментов, работа с фрагментами (копирование, удаление, перемещение).
- V. СПОСОБЫ ПОЛУЧЕНИЯ И ХИМИЧЕСКИЕ СВОЙСТВА АЦЕТИЛЕНОВЫХ И ДИЕНОВЫХ УГЛЕВОДОРОДОВ
- VII. Риски и способы их минимизации
- Yandex_direct_print() Теплоизоляция - требования, способы крепления
- Абсолютные и относительные показатели вариации и способы их расчёта.
- Абсолютные и средние показатели вариации и способы их расчета
- АДАПТАЦИЯ И ОСНОВНЫЕ СПОСОБЫ ПРИСПОСОБЛЕНИЯ ЖИВЫХ ОРГАНИЗМОВ К ЭКСТРЕМАЛЬНЫМ УСЛОВИЯМ СРЕДЫ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
