Сукцессии и дигрессии
Поступательные изменения в сообществе приводят в конечном счете к смене этого сообщества другим, с иным набором господствующих видов. Причиной подобных смен могут быть внешние по отношению к ценозу факторы, длительное время действующие в одном направлении, например возрастающее в результате мелиорации иссушение болотных почв, увеличивающееся загрязнение водоемов, усиленный выпас скота, вытаптывание лесопарков населением городов и т. п. Возникающие при этом смены одного биоценоза другим называют экзогенетическими. Если при этом усиливающееся влияние фактора приводит к постепенному упрощению структуры сообществ, обеднению их состава, снижению продуктивности, то подобные смены называют дигрессионными или дигрессиями.
Так, пастбищные дигрессии на террасовых песках Нижнего Днепра развиваются следующим образом. При умеренном выпасе степь находится на стадии дерновинных злаков. Преобладают типчак, кипец, житняк, ковыль. При дальнейшей усиленной пастьбе возникает стадия стержнекорневых двудольных с господством чаще всего молочая, а также двулетников и однолетников. Дерновинки злаков разбиваются копытами скота, а затем почти совершенно исчезают. На третьей стадии корневищных растений появляются злаки, характерные для сыпучих и слабозаросших песков: песчаный пырей, вейник, осока песчаная. На следующей стадии возникают голые пески с отдельными зарослями псаммофитов, характерных для предыдущего этапа. Таким образом, ковыльно-типчаковая степь сменяется сыпучими песками. Соответственно меняется и характер животного населения.
Эндогенетические смены возникают в результате процессов, происходящих внутри самого сообщества. Закономерный направленный процесс изменения сообществ в результате взаимодействия живых организмов между собой и окружающей их абиотической средой называют сукцессией.
Причины возникновения сукцессии.Сукцессия (от лат. successio– преемственность, наследование) – это процесс саморазвития сообществ. В основе сукцессии лежит неполнота биологического круговорота в данном ценозе. Каждый живой организм в результате жизнедеятельности меняет вокруг себя среду, изымая из нее часть веществ и насыщая ее продуктами метаболизма. При более или менее длительном существовании популяций они меняют свое окружение в неблагоприятную сторону и в результате оказываются вытесненными популяциями других видов, для которых вызванные преобразования среды оказываются экологически выгодными. Таким образом, в сообществе происходит смена господствующих видов. Длительное существование биоценоза возможно лишь в том случае, если изменения среды, вызванные деятельностью одних организмов, точно компенсируются деятельностью других, с противоположными экологическими требованиями.
В ходе сукцессии на основе конкурентных взаимодействий видов происходит постепенное формирование более устойчивых комбинаций, соответствующих конкретным абиотическим условиям среды. Примерами сукцессий могут быть смены видов при зарастании стоячих водоемов (рис. 153) или барханных песков в пустыне.
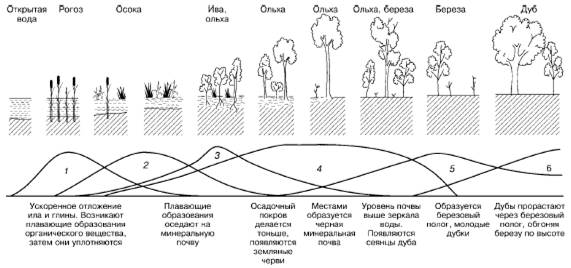
Рис. 153. Изменение экосистемы в ходе сукцессии при зарастании озера (по Д. Казенс, 1982): 1–6– смена доминирующих видов растений
Сыпучие барханные пески Каракумов и других районов Средней Азии сначала полностью лишены растительности и постоянного населения животных. Первым на них поселяется многолетний злак аристида, хорошо приспособленный к жизни в условиях постоянного переноса песка ветром. Корни у этого растения шнуровидные, и каждый заключен в чехол из сцементированных песчинок, что защищает корни от высыхания и механического повреждения, если они окажутся на поверхности. Побеги растут вертикально вверх и образуют дополнительные зоны кущения выше материнского, если песок засыпает растение. За счет аристиды уже могут существовать некоторые насекомые, и поэтому на барханы начинают забегать в поисках пищи ящерицы рода Eremias. На слегка скрепленных корнями аристиды песках получает возможность поселиться длиннокорневищная песчаная осока. Она успешно борется с песком, быстро прорастая сквозь его наносы и пронизывая песок ветвящимися корневищами на глубине 1–5 см. Покров разрежен, так как для обеспечения влагой одного растения нужна значительная площадь. На скрепленных злаками и осокой песках поселяются затем кустарники джузгун и белый саксаул, а также другие травянистые растения, в основном эфемеры: злаки, крестоцветные, мотыльковые, бурачниковые и т. п. Вслед за растительностью появляются растительноядные млекопитающие: тонкопалый суслик, мохноногий тушканчик, полуденная песчанка. Увеличивается видовое разнообразие насекомых – кормовой базы ящериц: ушастой и песчаной круглоголовок, сетчатой ящурки, гекконов. Появляются птицы – саксаульная сойка, дрофа-красотка, затем змея и хищные млекопитающие. Закрепленные пески Средней Азии отличаются большим видовым богатством и разнообразием жизненных форм растений и животных, так как водный режим их достаточно благоприятен: пески обладают способностью сгущать водяные пары в почве во влагу.
Последовательный ряд постепенно и закономерно сменяющих друг друга в сукцессии сообществ называется сукцессионной серией.
Сукцессии в природе чрезвычайно разномасштабны. Их можно наблюдать в пробирках с сенным настоем, где сменяют друг друга разные виды микроорганизмов и простейших (рис. 154), в лужах и прудах, в одной и той же порции листового опада в лесу, где происходят закономерные смены сапрофитов и сапрофагов, на стволах отмирающих деревьев, в пнях, на кротовинах, выбросах из нор сурков, зарастающих отмелях, выветривающихся скалах, на заброшенных пашнях, насыпях, на болотах, лугах, в лесах и т. д. Иерархичность в организации сообществ проявляется и в иерархичности сукцессионных процессов: более крупные преобразования биоценозов складываются из более мелких. Даже в стабильных экосистемах с хорошо отрегулированным круговоротом веществ постоянно осуществляется множество локальных сукцессионных смен, поддерживающих сложную внутреннюю структуру сообществ.
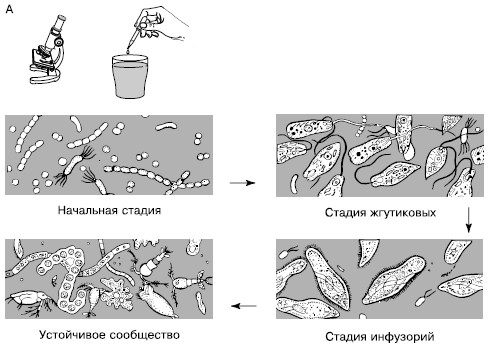
Рис. 154. Смена простейших в сенном настое (по Р. Дажо, 1975):
А – общая схема;
Б – конкретная серия:
1 – жгутиковые; 2 – инфузории кольноды; 3 – нарамеции; 4 – брюхо-ресничные инфузории; 5 – амебы; 6 – сувойки
Типы сукцессионных смен.Выделяют два основных типа сукцессионных смен: 1) с участием как автотрофного, так и гетеротрофного населения и 2) с участием лишь гетеротрофов. Сукцессии второго типа совершаются только в таких условиях, где создается предварительный запас или постоянное поступление органических соединений, за счет которых существует сообщество: в сильно загрязненных органическими веществами водоемах, в скоплениях разлагающейся растительной массы, в кучках или буртах навоза, компостах, в пещерах с гуано летучих мышей и т. п. (рис. 155).
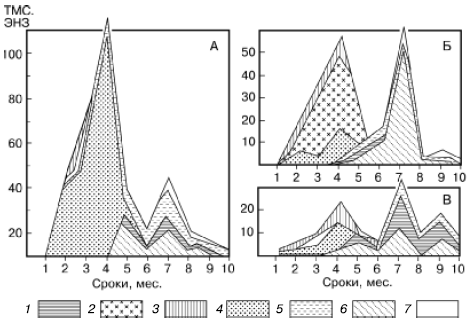
Рис. 155. Изменение численности разных групп микроартропод в ходе сукцессии при разложении растительных остатков клевера (А), картофельной ботвы (Б), ячменя (В) в пересчете на 100 г сухого органического вещества (по Н. М. Черновой, 1977): 1 – панцирные клещи; 2 – клещи-аноетиды; 3 – гипопусы аноетид; 4 – тироглифоидные клещи; 5 – их гипопусы; 6 – коллемболы; 7 – прочие группы
Сукцессии со сменой растительности могут быть первичными и вторичными.
Первичные сукцессии начинаются на лишенных жизни местах – на скалах, обрывах, наносах рек, сыпучих песках и т. п. При заселении таких участков живые организмы необратимо меняют местообитание и сменяют друг друга. Основная роль принадлежит накоплению отмерших растительных остатков или продуктов разложения, что зависит как от характера растительности, так и от комплекса разрушителей мертвой растительной массы – животных, грибов и микроорганизмов. Постепенно формируется почвенный профиль, изменяется гидрологический режим участка, его микроклимат. Такие сукцессии в геоботанике называют экогенетическими, так как они ведут к преобразованию самого местообитания.
В первичных сукцессиях, протекающих на скалах в лесах Урала, различают четыре этапа.
1. Поселение эндолитических и накипных лишайников, которые сплошь покрывают каменистую поверхность. Накипные лишайники несут своеобразную микрофлору и содержат богатую фауну простейших, коловраток и нематод. Мелкие клещи-сапрофаги и первичнобескрылые насекомые обнаруживаются сначала только в трещинах. Активность всего населения прерывиста, в основном после смачивания скал дождем или туманом.
2. Преобладание листоватых лишайников, постепенно образующих сплошной ковер. Под круговинками лишайников Parmelia в результате действия выделяемых ими кислот и механического сокращения слоевищ при высыхании образуются выщербленности, идет отмирание слоевищ и накопление детрита. Под лишайниками в большом количестве обитают мелкие членистоногие: коллемболы, панцирные клещи, сеноеды, личинки комаров-толкунчиков и другие, образуется микрогоризонт, состоящий из их экскрементов.
3. Поселения литофильных мхов Hedwigia ciliata и Pleurozium schreberi. Они погребают лишайники и подлишайниковые пленочные почвы. Ризоиды мхов прикрепляются не к камню, а к мелкозему, мощность которого достигает уже 3 см. Под мхами колебания температуры и влажности уже в несколько раз меньше, чем под лишайниками. Усиливается деятельность микроорганизмов, и увеличивается разнообразие групп животных.
4. Появление гипновых мхов и сосудистых растений. В разложении растительных остатков и формировании почвенного профиля постепенно уменьшается роль мелких членистоногих и растет участие более крупных беспозвоночных-сапрофагов: энхитреид, дождевых червей, личинок насекомых.
Вторичные сукцессии представляют собой восстановительные смены. Они начинаются в том случае, если в уже сложившихся сообществах частично нарушены установившиеся взаимосвязи организмов, например удалена растительность одного или нескольких ярусов (в результате вырубки, пожара, вспашки и т. д.). Смены, ведущие к восстановлению прежнего состава ценоза, получили в геоботанике название демутационных. Примером может служить демутация залежей в Абаканских степях, происходящая в четыре основных этапа: 1) преобладание однолетних сорняков – на 1-2-й год после вспашки; 2) господство «крупного бурьяна», преимущественно разных полыней – на 3-4-й год; 3) вытеснение их корневищными злаками (пыреем и др.) и появление большого количества бобовых – начиная с 5-го года; 4) установление господства степных дерновинных злаков с появлением ковыля – на 11-12-й год после вспашки.
Восстановительные смены совершаются быстрее и легче, чем экогенетические, так как в нарушенном местообитании сохраняются почвенный профиль, семена, зачатки и часть прежнего населения и прежних связей. Демутации не являются повторением какого-либо этапа первичных сукцессии.
Частными вариантами сукцессии являются смены, которые по продолжительности точно совпадают с каким-либо сезоном, а на следующий год начинаются сначала. Эти «сезонные» сукцессии выглядят как обычная фенологическая смена форм, но природа их иная. Например, в планктоне северных морей весной массовое размножение водорослей сменяется волной размножения рачков-калянусов, затем в большом количестве появляются гребневики, а после них – некоторые виды рыб. Эти смены происходят как постепенное развертывание в течение короткого северного лета единой гигантской трофической цепи организмов, так как каждая последующая группа питается за счет предыдущей. Если же в силу неблагоприятной ситуации не возникает, например, весной массовое размножение фитопланктона, то и все последующие группы будут угнетены, несмотря на благоприятные сочетания любых прочих факторов в период обычного появления группы.
Своеобразный вариант смен представляют поточные, или конвейерные, сукцессии. Они развиваются в подвижной среде: реках, потоках, круговоротах водных масс в океанах. Планктонные сообщества, увлекаемые водой, на разных стадиях развития оказываются в разных географических точках, а в одном и том же регионе постоянны по составу. Такой же конвейерный тип преобразований наблюдается, например, в лесной подстилке (рис. 156). В разных ее подгоризонтах: верхнем рыхлом слое опада, среднем спрессованном «ферментативном» подгоризонте и нижнем, полностью переработанном муллевом слое – существует постоянная значительная разница в многочисленном микронаселении. Но в каждой конкретной порции опада, поступающего сверху, происходит сукцессионная смена микроорганизмов, простейших, нематод и мелких членистоногих, обеспечивающая все более глубокое разложение растительных остатков. Опад вместе со всем населением постепенно занимает все более глубокое положение, захораниваясь под новыми поступлениями, а общая стратификация подстилки выглядит неизменной.
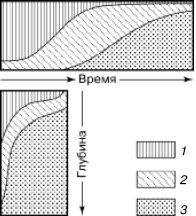
Рис. 156. Аналогии в сменах жизненных форм мелких членистоногих в изолированном опаде (вверху) и верхних слоях почвенного профиля (внизу) (по Н. М. Черновой, 1977):
1– поверхностные формы микроартропод;
2– полупочвенные микроартроподы;
3– почвенные формы
Процесс сукцессии.Процесс сукцессии, по Ф. Клементсу, состоит из нескольких этапов: 1) возникновения незанятого жизнью участка; 2) миграции на него различных организмов или их зачатков; 3) приживания их на данном участке; 4) конкуренции их между собой и вытеснения отдельных видов; 5) преобразования живыми организмами местообитания, постепенной стабилизации условий и отношений.
В настоящее время практически вся доступная жизни поверхность суши занята различными сообществами, и поэтому возникновение свободных от живых существ участков имеет локальный характер. Это или места, освободившиеся в результате отодвигания ледников, отступания уреза воды в водоемах, обвалов, эрозии и т. п., или возникшие в результате деятельности человека, например выноса наверх больших масс глубинных пород при разработке полезных ископаемых.
Занос спор, семян растений, проникновение животных на освободившийся участок имеют большей частью случайный характер и зависят от того, какие виды есть в окружающих биотопах. Из числа видов, попавших в новое местообитание, приживаются лишь те, экологическая валентность которых соответствует данному комплексу абиотических условий. Прижившиеся виды постепенно занимают весь новый биотоп, вступая в конкуренцию друг с другом. В результате происходит перестройка видового состава и количественных соотношений разных форм. Параллельно идет процесс преобразования самого местообитания под влиянием развивающегося сообщества. Процесс завершается формированием более или менее стабильной системы с уравновешенным типом биологического круговорота.
Сукцессии любого масштаба и ранга характеризуются целым рядом общих закономерностей, многие из которых чрезвычайно важны для практической деятельности человека.
В любой сукцессионной серии темпы происходящих изменений постепенно замедляются. Конечным итогом является формирование относительно устойчивой стадии – климаксового сообщества, или климакса. Начальные, пионерные группировки видов отличаются наибольшей динамичностью и неустойчивостью. Климаксовые же экосистемы способны к длительному самоподдерживанию в соответствующем диапазоне условий, так как приобретают такие черты организации биоценозов, которые позволяют поддерживать сбалансированный круговорот веществ.
В ходе сукцессии постепенно нарастает видовое многообразие. Это ведет к усложнению связей внутри ценоза, разветвлению цепей питания и усложнению трофической сети, умножению симбиотических отношений, усилению регуляторных возможностей внутри системы. Тем самым уменьшается вероятность слишком сильного размножения отдельных видов и снижается степень доминирования наиболее массовых форм.
Чрезвычайное увеличение численности отдельных видов возможно чаще всего на начальных этапах развития сукцессионных серий, когда еще в недостаточной мере сложилась система взаимного регулирования. В незрелых сообществах, т. е. находящихся в начале сукцессионных рядов, преобладают мелкоразмерные виды с короткими жизненными циклами и высоким потенциалом размножения, специализированные на быстром захвате освободившегося пространства. Они обычно обладают широкими расселительными возможностями, позволяющими им первыми проникать на незанятые участки, но малоспособны к конкуренции и длительному удержанию за собой пространства.
Постепенно в развивающихся сообществах появляются и закрепляются более крупные формы с длительными и сложными циклами развития. Нарастание экологического разнообразия ведет к более четкому распределению групп организмов по экологическим нишам. В растительном покрове становится сильнее выражена ярусность и мозаичность, создающие основу пространственной структуры наземных экосистем. Усиливается зависимость успешного существования одних видов от биохимических выделений, роста или поведения других: преобладание конкурентных взаимоотношений сменяется преобладанием мутуалистических и трофических зависимостей.
В результате сообщества приобретают известную степень автономности и независимости от окружающих условий, не подчиняя свою жизнь флюктуациям внешней среды, а вырабатывая собственные эндогенные ритмы.
Не меньшие преобразования происходят и в энергетическом балансе системы (рис. 157). С энергетических позиций сукцессия – такое неустойчивое состояние сообщества, которое характеризуется неравенством двух показателей: общей продуктивности и энергетических трат всей системы на поддержание обмена веществ.
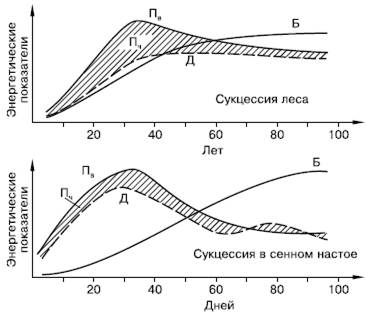
Рис. 157. Изменение энергетических показателей сообществ при развитии соснового леса (вверху) и лабораторной культуры простейших в сенном настое (внизу) (по Ю. Одуму, 1975):
Пв – продукция валовая; Д – траты на дыхание; Пч – продукция чистая (заштриховано); Б – биомасса
В ходе сукцессии общая биомасса сообщества сначала возрастает, но затем темпы этого прироста снижаются, и на стадии климакса биомасса системы стабилизируется. Это происходит потому, что на первых этапах сукцессии, когда видовой состав сообществ еще беден и пищевые цепи коротки, не вся часть прироста растительной массы потребляется гетеротрофами. Таким образом, относительно высока чистая продукция сообщества, которая идет на увеличение его биомассы. Накапливаются как общая масса живых организмов, так и запасы мертвого, неразложившегося органического вещества. В зрелых, устойчивых экосистемах практически весь годовой прирост растительности поступает и расходуется в цепях питания гетеротрофами, поэтому чистая продукция биоценоза, его «урожай», приближается к нулю.
Биогенные элементы растения сначала черпают из запасов почвы, но постепенно, по мере истощения этих запасов и накопления в системе мертвого органического вещества, разложение его становится основным источником минерального питания растений, и круговороты биогенных веществ из открытых превращаются в закрытые.
Неравенство трат энергии на образование первичной продукции и суммарный метаболизм (дыхание) сообщества проявляется в тех сукцессиях, в которых участвуют только гетеротрофные организмы. В этом частном случае первичная продукция равна нулю, и если приток мертвого органического вещества, за счет которого существует сообщество, не восполняет запасы в системе, то сообщество вскоре истощает свою энергетическую базу.
С удлинением цепей питания увеличивается эффективность использования поступающей энергии, иначе – КПД всей системы, так как одна и та же порция энергии идет на поддержание большого количества биомассы. Знание этих закономерностей имеет большое значение в практической деятельности человека.
Изымая избыток чистой продукции из биоценозов, находящихся в начале сукцессионных рядов, мы задерживаем сукцессию, но не подрываем основу существования сообщества. Вмешательство же в стабильные, климаксовые системы, с большой полнотой расходующие энергию на «свои» нужды, неминуемо вызывает нарушения сложившегося равновесия. Пока нарушения не превышают самовосстановительной способности ценоза, возникающие демутационные смены могут вернуть его к исходному состоянию. Этим пользуются, например, при рациональном планировании рубок леса. Но если сила и частота воздействия выходят за рамки этих возможностей, то первоначально устойчивое, богатое видами сообщество постепенно деградирует, сменяясь производными с малой способностью к самовозобновлению.
Вырубка леса на локальных участках с оставлением части территории под коренными типами лесной растительности вызывает ускоренные сукцессии, исходные фитоценозы восстанавливаются за относительно короткий срок – немногие десятилетия. Сплошные же рубки на больших площадях, особенно если используется мощная корчевальная техника, полностью разрушают не только лесное сообщество, но и весь почвенный покров, эволюция которого шла тысячелетия. В этом случае сукцессии приводят к иным, упрощенным типам сообществ, и на месте лесов возникают пустоши, болота или другие малопродуктивные экосистемы.
Таким образом, сообщество не может одновременно сочетать два противоположных свойства: быть высокостабильным и давать большой запас чистой продукции, который можно было бы изымать без вреда для самого ценоза.
Сукцессии, протекающие при разложении органического вещества в почве, лежат в основе биологического круговорота. Это естественные регуляторные процессы, восстанавливающие нормальное состояние нарушенных почв. Такие проблемы века, как подрыв естественного плодородия в результате нарушения процессов образования гумуса, загрязнение среды ядохимикатами и органическими отходами, эрозия, «утомление почв» и другие отрицательные явления, возникли вследствие ослабления регуляторных возможностей почв.
Стабильность экосистем, надежность протекания биологического круговорота веществ основаны на видовом разнообразии и полноте сукцессий.
Агроэкосистемы
Агроэкосистемы (сельскохозяйственные экосистемы), создаваемые человеком для получения высокой чистой продукции автотрофов (урожая), отличаются от природных рядом особенностей:
1. В них резко снижено разнообразие организмов. На полях обычно культивируют один или немного видов растений, в связи с чем резко обедняется и животное население, и состав микроорганизмов в биоценозе. Выпас животных также сильно упрощает видовую структуру пастбищных сообществ. Культурные пастбища с подсевом трав приближаются по этому показателю к полям сельскохозяйственных растений. Видовое разнообразие разводимых человеком животных ничтожно мало по сравнению с природным.
2. Виды, культивируемые человеком, поддерживаются искусственным отбором в состоянии, далеком от первоначального, и не могут выдерживать борьбу за существование с дикими видами без поддержки человека.
3. Агроэкосистемы получают дополнительный поток энергии, кроме солнечной, благодаря деятельности людей, животных и механизмов, обеспечивающих необходимые условия роста культивируемых видов. Чистая первичная продукция (урожай) удаляется из экосистемы и не поступает в цепи питания. Частичное использование ее вредителями представляет нежелательное явление и всячески пресекается деятельностью человека.
В настоящее время пахотными землями и пастбищами занято свыше 30 % суши, и деятельность людей по поддержанию этих систем превращается в глобальный экологический фактор.
Несмотря на значительную упрощенность агроэкосистем, в них все же сохраняется множество биоценотических связей, в конечном счете влияющих на судьбу урожая (рис. 158). Сопоставление сведений о фауне и флоре пшеничных полей показывает гигантскую сложность даже предельно простого агроценоза, здесь сохраняется более тысячи видов.
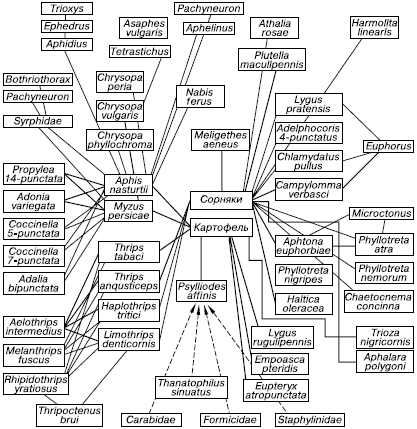
Рис. 158. Биоценотические связи на картофельном поле (по В. Тишлеру,1971): сплошные линии – виды, обитающие на растениях; пунктирные – виды, обитающие на поверхности почвы; утолщенные линии – доминирующие виды
История формирования ценотических группировок, связанных с основными сельскохозяйственными культурами, насчитывает немногие сотни лет. Буквально на глазах человека формируются сообщества, приспособленные к жесткому прессу агротехнических режимов, с широким размахом колебательных циклов в жизни популяций, с четко отобранным кругом доминантов. Наглядно проявляются эволюционные сдвиги в экологических характеристиках и адаптивных показателях разных видов.
Условия, которым в идеале должны соответствовать поля сельскохозяйственных культур, – быть высокопродуктивными и вместе с тем стабильными – с экологической точки зрения несовместимы. В природных экосистемах первичная продукция растений потребляется в многочисленных цепях питания и вновь возвращается в виде минеральных солей и углекислого газа в систему биологического круговорота. Ограждая урожай от его природных потребителей, отчуждая его и заменяя естественный опад органическими и минеральными удобрениями, мы обрываем множество цепей питания и дисбалансируем сообщество. По существу, все усилия по созданию высокой чистой продукции отдельных культур в пользу человека есть борьба «против природы», которая требует большой затраты труда и материальных средств.
Вместе с тем агроценозы выступают гигантской лабораторией, где человек учится, используя отдельные звенья системы, управлять продукционным процессом и круговоротом веществ, регулировать численность популяций. Тщательное изучение таких относительно упрощенных систем, как агроценозы, вносит серьезный вклад в развитие общей биоценологии. В обедненных сообществах более резко вырисовываются те законы, которые лежат в основе объединения живых существ в надорганизменные системы. Богатый арсенал агротехнических средств, имеющийся на вооружении современного сельского хозяйства, позволяет экспериментально и в широких масштабах проверять различные пути воздействия на сообщества, оценивать степень их устойчивости и прочность связей в отдельных звеньях.
Все искусственно создаваемые в сельскохозяйственной практике экосистемы полей, садов, пастбищных лугов, огородов, теплиц и других агроценозов представляют собой системы, специально поддерживаемые человеком на начальных стадиях сукцессионных преобразований. В агроценозах используется именно свойство пионерных сообществ производить высокую чистую продукцию. Но такие сообщества и в природе неустойчивы, не способны к самовозобновлению и саморегулированию, подвержены угрозе гибели от массового размножения вредителей или болезней (рис. 159). Они требуют неустанной деятельности по их поддержанию со стороны человека.
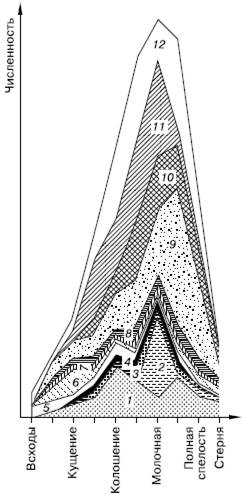
Рис. 159. Динамика численности насекомых в стеблестое овса (по Н. И. Куликову, 1985): 1– цикадки; 2 – их личинки; 3 – хлебный клопик; 4 – клопы-лигулы; 5 – хлебная полосатая блошка; 6 – жуки-блестянки; 7 – божьи коровки; 8 – паразитические перепончатокрылые; 9 – шведская муха; 10 – злаковые трипсы; 11 – злаковые тли; 12 – общая численность
Сельскохозяйственное освоение территорий часто приводит к разрушению созданных природой механизмов регуляции численности отдельных видов и резким изменениям в уровне их обилия.
В агроценозах чаще всего происходят «экологические взрывы», как назвал Ч. Элтон чрезмерное увеличение численности отдельных видов. Последствия этих «взрывов» могут быть весьма существенными для сельского и лесного хозяйства. Так, распространение патогенного грибка фитофторы из Европы в Ирландию в прошлом столетии погубило весь урожай картофеля, вызвав сильный голод. Во Франции более 1 млн га виноградников пришлось уничтожить в результате размножения корневой тли – филлоксеры.
Искусственная регуляция численности вредителей – по большей части необходимое условие поддержания агроэкосистем. Это связано прежде всего с необходимостью подавления видов, вышедших из-под контроля естественных регуляторных механизмов. В ряде случаев даже полное сохранение естественной регуляции численности вида не удовлетворяет экономическим требованиям. Например, стабилизация численности яблонной плодожорки на уровне, при котором погибает большая часть урожая, с позиций естественного отбора не угрожает существованию яблони как вида. С хозяйственной же точки зрения необходимо резкое понижение уровня, на котором должна находиться численность плодожорки в садах. Поэтому в сельскохозяйственной практике применяют мощные средства подавления численности нежелательных видов: ядохимикаты, гербициды и т. д. Экологические последствия этих действий приводят, однако, к ряду нежелательных эффектов, кроме тех, для которых они применяются.
Подавление численности вредителей химическими средствами, кроме загрязнения среды и включения ядов в цепи питания, часто вызывает так называемый «бумеранг-эффект»: вслед за подавлением численности вредителя вскоре возникает новая, еще более мощная его вспышка. Обычно применение ядохимикатов тотального действия сильнее влияет на естественных врагов вредителя, чем на его собственные популяции. В результате следующие поколения полностью освобождаются из-под пресса паразитов и хищников и осуществляется их массовое размножение. Таким образом, недоучет биоценотических механизмов регуляции численности на полях сельскохозяйственных культур также не в экономических интересах человека. В трехзвенной цепи: культурное растение – вредитель – паразит повышение чистой продукции растений может быть достигнуто как подавлением второго звена, так и усилением третьего. Именно этот подход используется в разработке биологических методов борьбы с вредителями.
В отношении к сообществам, складывающимся в агроэкосистемах, в связи с общим развитием экологических знаний постепенно меняются акценты. На смену представлениям об обрывочности, осколочности ценотических связей и предельной упрощенности агроценозов возникает понимание их сложной системной организации, где человек существенно влияет лишь на отдельные звенья, а вся система продолжает развиваться по естественным, природным законам (рис. 160).
Рис. 160. Фазовая приуроченность насекомых-вредителей на посевах озимой пшеницы (по Н. И. Куликову, 1985):
1– злаковые тли; 2 – цикадка шеститочечная; 3 – цикадка полосатая; 4 – клопы-лигусы; 5 – хлебный клопик, 6 – злаковые трипсы; 7 – хлебная полосатая блошка; 8 – ростковая муха; 9 – яровая муха; 10 – шведская муха
Современные представления плодотворны и в теоретическом, и в практическом плане. Они стимулируют поиск внутренних регуляторных резервов агроценозов, использование которых позволяет не допускать массового размножения вредителей и негативных последствий химической борьбы с ними. В создании оптимальной структуры культурных фитоценозов также кроются новые возможности увеличения пределов первичной продукции. Изучение основных законов организации и пределов стабильности агроценозов дает теоретической экологии уникальный материал.
С экологических позиций крайне опасно упрощать природное окружение человека, превращая весь ландшафт в агрохозяйственный. Основная стратегия по созданию высокопродуктивного и устойчивого ландшафта должна заключаться в сохранении и умножении его многообразия. «Если дикая природа отступает, – писал Ч. Элтон, – мы должны научиться передавать часть ее стойкости и богатства ландшафтам тех земель, с которых мы снимаем наши урожаи». Наряду с поддержанием высокопродуктивных полей следует особенно заботиться о сохранении как можно более многообразных заповедных, не подвергающихся усиленному антропогенному воздействию участков разного масштаба, с богатым видовым разнообразием, которые могли бы быть источником видов для восстанавливающихся в сукцессионных рядах сообществ. Эксплуатация ценных для человека природных систем не должна превышать их способности к самовосстановлению. Аграрный ландшафт должен быть разнообразным, с лесными полосами вокруг полей, живыми изгородями и перелесками.
Идея плановой реконструкции сельскохозяйственных ландшафтов родилась в нашей стране и связана с именами В. В. Докучаева и А. И. Воейкова. В настоящее время эти идеи приобретают особый экологический смысл, так как входят составной частью в общую стратегию охраны природы, окружающей человека.
Глава 10. БИОСФЕРА
Понятие о биосфере
Идея о влиянии жизни на природные процессы на огромных пространствах Земли была впервые научно обоснована на рубеже XIX и XX столетий в трудах В. В. Докучаева, который указал на зависимость типа почвообразования не только от климата, но и от совокупного влияния растительности и животных. Термин «биосфера» использовал в 1875 г. австрийский геолог Э. Зюсс для обозначения оболочки Земли, населяемой живыми организмами.
В 20-х годах прошедшего века в трудах В. И. Вернадского было разработано представление о биосфере как глобальной единой системе Земли, где весь основной ход геохимических и энергетических превращений определяется жизнью. Идеи В. И. Вернадского намного опередили состояние современной ему науки и в должной мере были оценены лишь во второй половине века, после возникновения концепции экосистем. Большинство процессов, меняющих в течение геологического времени лик нашей планеты, рассматривали ранее как чисто физические, химические или физико-химические явления (размыв, растворение, осаждение, гидролиз и т. п.). В. И. Вернадский впервые создал учение о геологической роли живых организмов, показав, что деятельность живых существ является главным фактором преобразования земной коры.
Биосферой В. И. Вернадский назвал ту область нашей планеты, в которой существует или когда-либо существовала жизнь и которая постоянно подвергается или подвергалась воздействию живых организмов.
Участие каждого отдельного организма в геологической истории Земли ничтожно мало. Однако живых существ на Земле бесконечно много, они обладают высоким потенциалом размножения, активно взаимодействуют со средой обитания и в конечном счете представляют в своей совокупности особый, глобальных масштабов фактор, преобразующий верхние оболочки Земли.
Значение организмов обусловлено их большим разнообразием, повсеместным распространением, длительностью существования в истории Земли, избирательным характером биохимической деятельности и исключительно высокой химической активностью по сравнению с другими компонентами природы.
Всю совокупность организмов на планете В. И. Вернадский назвал живым веществом, рассматривая в качестве его основных характеристик суммарную массу, химический состав и энергию.
Косное вещество, по В. И. Вернадскому, – это совокупность тех веществ в биосфере, в образовании которых живые организмы не участвуют.
Биогенное вещество создается и перерабатывается жизнью, совокупностями живых организмов. Это источник чрезвычайно мощной потенциальной энергии (каменный уголь, битумы, известняки, нефть). После образования биогенного ве
Дата добавления: 2016-07-11; просмотров: 3434;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
