Эндоплазматическая сеть
Эндоплазматическая сеть (ЭПС), или эндоплазматический ретикулум (ЭПР), — одномембранный органоид. Представляет собой систему мембран, формирующих «цистерны» и каналы, соединенных друг с другом и ограничивающих единое внутреннее пространство — полости ЭПС. Мембраны с одной стороны связаны с цитоплазматической мембраной, с другой — с наружной ядерной мембраной. Различают два вида ЭПС: 1) шероховатая (гранулярная), содержащая на своей поверхности рибосомы, и 2) гладкая (агранулярная), мембраны которой рибосом не несут.
Функции: 1) транспорт веществ из одной части клетки в другую, 2) разделение цитоплазмы клетки на компартменты ( «отсеки»), 3) синтез углеводов и липидов (гладкая ЭПС), 4) синтез белка (шероховатая ЭПС), 5) место образования аппарата Гольджи.
Аппарат Гольджи
Аппарат Гольджи, или комплекс Гольджи, — одномембранный органоид. Представляет собой стопки уплощенных «цистерн» с расширенными краями. С ними связана система мелких одномембранных пузырьков (пузырьки Гольджи). Каждая стопка обычно состоит из 4-х–6-ти «цистерн», является структурно-функциональной единицей аппарата Гольджи и называется диктиосомой. Число диктиосом в клетке колеблется от одной до нескольких сотен. В растительных клетках диктиосомы обособлены.

Аппарат Гольджи обычно расположен около клеточного ядра (в животных клетках часто вблизи клеточного центра).
Функции аппарата Гольджи: 1) накопление белков, липидов, углеводов, 2) модификация поступивших органических веществ, 3) «упаковка» в мембранные пузырьки белков, липидов, углеводов, 4) секреция белков, липидов, углеводов, 5) синтез углеводов и липидов, 6) место образования лизосом. Секреторная функция является важнейшей, поэтому аппарат Гольджи хорошо развит в секреторных клетках.
Лизосомы
Лизосомы — одномембранные органоиды. Представляют собой мелкие пузырьки (диаметр от 0,2 до 0,8 мкм), содержащие набор гидролитических ферментов. Ферменты синтезируются на шероховатой ЭПС, перемещаются в аппарат Гольджи, где происходит их модификация и упаковка в мембранные пузырьки, которые после отделения от аппарата Гольджи становятся собственно лизосомами. Лизосома может содержать от 20 до 60 различных видов гидролитических ферментов. Расщепление веществ с помощью ферментов называют лизисом.
Различают: 1) первичные лизосомы, 2) вторичные лизосомы. Первичными называются лизосомы, отшнуровавшиеся от аппарата Гольджи. Первичные лизосомы являются фактором, обеспечивающим экзоцитоз ферментов из клетки.
Вторичными называются лизосомы, образовавшиеся в результате слияния первичных лизосом с эндоцитозными вакуолями. В этом случае в них происходит переваривание веществ, поступивших в клетку путем фагоцитоза или пиноцитоза, поэтому их можно назвать пищеварительными вакуолями.
Яндекс.ДиректВсе объявления
Автофагия — процесс уничтожения ненужных клетке структур. Сначала подлежащая уничтожению структура окружается одинарной мембраной, затем образовавшаяся мембранная капсула сливается с первичной лизосомой, в результате также образуется вторичная лизосома (автофагическая вакуоль), в которой эта структура переваривается. Продукты переваривания усваиваются цитоплазмой клетки, но часть материала так и остается непереваренной. Вторичная лизосома, содержащая этот непереваренный материал, называется остаточным тельцем. Путем экзоцитоза непереваренные частицы удаляются из клетки.
Автолиз — саморазрушение клетки, наступающее вследствие высвобождения содержимого лизосом. В норме автолиз имеет место при метаморфозах (исчезновение хвоста у головастика лягушек), инволюции матки после родов, в очагах омертвления тканей.
Функции лизосом: 1) внутриклеточное переваривание органических веществ, 2) уничтожение ненужных клеточных и неклеточных структур, 3) участие в процессах реорганизации клеток.
Вакуоли
Вакуоли — одномембранные органоиды, представляют собой «емкости», заполненные водными растворами органических и неорганических веществ. В образовании вакуолей принимают участие ЭПС и аппарат Гольджи. Молодые растительные клетки содержат много мелких вакуолей, которые затем по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну большую центральную вакуоль. Центральная вакуоль может занимать до 95% объема зрелой клетки, ядро и органоиды оттесняются при этом к клеточной оболочке. Мембрана, ограничивающая растительную вакуоль, называется тонопластом. Жидкость, заполняющая растительную вакуоль, называется клеточным соком. В состав клеточного сока входят водорастворимые органические и неорганические соли, моносахариды, дисахариды, аминокислоты, конечные или токсические продукты обмена веществ (гликозиды, алкалоиды), некоторые пигменты (антоцианы).
В животных клетках имеются мелкие пищеварительные и автофагические вакуоли, относящиеся к группе вторичных лизосом и содержащие гидролитические ферменты. У одноклеточных животных есть еще сократительные вакуоли, выполняющие функцию осморегуляции и выделения.
Функции вакуоли: 1) накопление и хранение воды, 2) регуляция водно-солевого обмена, 3) поддержание тургорного давления, 4) накопление водорастворимых метаболитов, запасных питательных веществ, 5) окрашивание цветов и плодов и привлечение тем самым опылителей и распространителей семян, 6) см. функции лизосом.
Эндоплазматическая сеть, аппарат Гольджи, лизосомы и вакуоли образуют единую вакуолярную сеть клетки, отдельные элементы которой могут переходить друг в друга.
Митохондрии
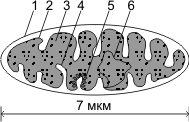
Строение митохондрии:
1 — наружная мембрана;
2 — внутренняя мембрана; 3 — матрикс; 4 — криста; 5 — мультиферментная система; 6 — кольцевая ДНК.
Форма, размеры и количество митохондрий чрезвычайно варьируют. По форме митохондрии могут быть палочковидными, округлыми, спиральными, чашевидными, разветвленными. Длина митохондрий колеблется в пределах от 1,5 до 10 мкм, диаметр — от 0,25 до 1,00 мкм. Количество митохондрий в клетке может достигать нескольких тысяч и зависит от метаболической активности клетки.
Митохондрия ограничена двумя мембранами. Наружная мембрана митохондрий (1) гладкая, внутренняя (2) образует многочисленные складки — кристы (4). Кристы увеличивают площадь поверхности внутренней мембраны, на которой размещаются мультиферментные системы (5), участвующие в процессах синтеза молекул АТФ. Внутреннее пространство митохондрий заполнено матриксом (3). В матриксе содержатся кольцевая ДНК (6), специфические иРНК, рибосомы прокариотического типа (70S-типа), ферменты цикла Кребса.
Митохондриальная ДНК не связана с белками («голая»), прикреплена к внутренней мембране митохондрии и несет информацию о строении примерно 30 белков. Для построения митохондрии требуется гораздо больше белков, поэтому информация о большинстве митохондриальных белков содержится в ядерной ДНК, и эти белки синтезируются в цитоплазме клетки. Митохондрии способны автономно размножаться путем деления надвое. Между наружной и внутренней мембранами находится протонный резервуар, где происходит накопление Н+.
Функции митохондрий: 1) синтез АТФ, 2) кислородное расщепление органических веществ.
Согласно одной из гипотез (теория симбиогенеза) митохондрии произошли от древних свободноживущих аэробных прокариотических организмов, которые, случайно проникнув в клетку-хозяина, затем образовали с ней взаимовыгодный симбиотический комплекс. В пользу этой гипотезы свидетельствуют следующие данные. Во-первых, митохондриальная ДНК имеет такие же особенности строения как и ДНК современных бактерий (замкнута в кольцо, не связана с белками). Во-вторых, митохондриальные рибосомы и рибосомы бактерий относятся к одному типу — 70S-типу. В-третьих, механизм деления митохондрий сходен с таковым бактерий. В-четвертых, синтез митохондриальных и бактериальных белков подавляется одинаковыми антибиотиками.
Пластиды
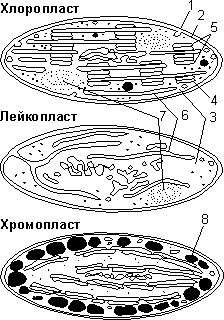
Строение пластид: 1 — наружная мембрана; 2 — внутренняя мембрана; 3 — строма; 4 — тилакоид; 5 — грана; 6 — ламеллы; 7 — зерна крахмала; 8 — липидные капли.
Пластиды характерны только для растительных клеток. Различают три основных типа пластид: лейкопласты — бесцветные пластиды в клетках неокрашенных частей растений, хромопласты — окрашенные пластиды обычно желтого, красного и оранжевого цветов, хлоропласты — зеленые пластиды.
Хлоропласты. В клетках высших растений хлоропласты имеют форму двояковыпуклой линзы. Длина хлоропластов колеблется в пределах от 5 до 10 мкм, диаметр — от 2 до 4 мкм. Хлоропласты ограничены двумя мембранами. Наружная мембрана (1) гладкая, внутренняя (2) имеет сложную складчатую структуру. Наименьшая складка называется тилакоидом (4). Группа тилакоидов, уложенных наподобие стопки монет, называется граной (5). В хлоропласте содержится в среднем 40–60 гран, расположенных в шахматном порядке. Граны связываются друг с другом уплощенными каналами — ламеллами (6). В мембраны тилакоидов встроены фотосинтетические пигменты и ферменты, обеспечивающие синтез АТФ. Главным фотосинтетическим пигментом является хлорофилл, который и обусловливает зеленый цвет хлоропластов.
Внутреннее пространство хлоропластов заполнено стромой (3). В строме имеются кольцевая «голая» ДНК, рибосомы 70S-типа, ферменты цикла Кальвина, зерна крахмала (7). Внутри каждого тилакоида находится протонный резервуар, происходит накопление Н+. Хлоропласты, также как митохондрии, способны к автономному размножению путем деления надвое. Они содержатся в клетках зеленых частей высших растений, особенно много хлоропластов в листьях и зеленых плодах. Хлоропласты низших растений называют хроматофорами.
Функция хлоропластов: фотосинтез. Полагают, что хлоропласты произошли от древних эндосимбиотических цианобактерий (теория симбиогенеза). Основанием для такого предположения является сходство хлоропластов и современных бактерий по ряду признаков (кольцевая, «голая» ДНК, рибосомы 70S-типа, способ размножения).
Лейкопласты. Форма варьирует (шаровидные, округлые, чашевидные и др.). Лейкопласты ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя образует малочисленные тилакоиды. В строме имеются кольцевая «голая» ДНК, рибосомы 70S-типа, ферменты синтеза и гидролиза запасных питательных веществ. Пигменты отсутствуют. Особенно много лейкопластов имеют клетки подземных органов растения (корни, клубни, корневища и др.). Функция лейкопластов: синтез, накопление и хранение запасных питательных веществ. Амилопласты — лейкопласты, которые синтезируют и накапливают крахмал, элайопласты — масла, протеинопласты — белки. В одном и том же лейкопласте могут накапливаться разные вещества.
Хромопласты. Ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя или также гладкая, или образует единичные тилакоиды. В строме имеются кольцевая ДНК и пигменты — каротиноиды, придающие хромопластам желтую, красную или оранжевую окраску. Форма накопления пигментов различная: в виде кристаллов, растворены в липидных каплях (8) и др. Содержатся в клетках зрелых плодов, лепестков, осенних листьев, редко — корнеплодов. Хромопласты считаются конечной стадией развития пластид.
Функция хромопластов: окрашивание цветов и плодов и тем самым привлечение опылителей и распространителей семян.
Все виды пластид могут образовываться из пропластид. Пропластиды — мелкие органоиды, содержащиеся в меристематических тканях. Поскольку пластиды имеют общее происхождение, между ними возможны взаимопревращения. Лейкопласты могут превращаться в хлоропласты (позеленение клубней картофеля на свету), хлоропласты — в хромопласты (пожелтение листьев и покраснение плодов). Превращение хромопластов в лейкопласты или хлоропласты считается невозможным.
Рибосомы
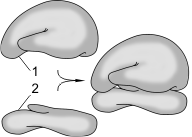
Строение рибосомы:
1 — большая субъединица; 2 — малая субъединица.
Рибосомы — немембранные органоиды, диаметр примерно 20 нм. Рибосомы состоят из двух субъединиц — большой и малой, на которые могут диссоциировать. Химический состав рибосом — белки и рРНК. Молекулы рРНК составляют 50–63% массы рибосомы и образуют ее структурный каркас. Различают два типа рибосом: 1) эукариотические (с константами седиментации целой рибосомы — 80S, малой субъединицы — 40S, большой — 60S) и 2) прокариотические (соответственно 70S, 30S, 50S).
В составе рибосом эукариотического типа 4 молекулы рРНК и около 100 молекул белка, прокариотического типа — 3 молекулы рРНК и около 55 молекул белка. Во время биосинтеза белка рибосомы могут «работать» поодиночке или объединяться в комплексы — полирибосомы (полисомы). В таких комплексах они связаны друг с другом одной молекулой иРНК. Прокариотические клетки имеют рибосомы только 70S-типа. Эукариотические клетки имеют рибосомы как 80S-типа (шероховатые мембраны ЭПС, цитоплазма), так и 70S-типа (митохондрии, хлоропласты).
Субъединицы рибосомы эукариот образуются в ядрышке. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка.
Функция рибосом: сборка полипептидной цепочки (синтез белка).
Цитоскелет
Цитоскелет образован микротрубочками и микрофиламентами. Микротрубочки — цилиндрические неразветвленные структуры. Длина микротрубочек колеблется от 100 мкм до 1 мм, диаметр составляет примерно 24 нм, толщина стенки — 5 нм. Основной химический компонент — белок тубулин. Микротрубочки разрушаются под воздействием колхицина. Микрофиламенты — нити диаметром 5–7 нм, состоят из белка актина. Микротрубочки и микрофиламенты образуют в цитоплазме сложные переплетения. Функции цитоскелета: 1) определение формы клетки, 2) опора для органоидов, 3) образование веретена деления, 4) участие в движениях клетки, 5) организация тока цитоплазмы.
Клеточный центр
Клеточный центр включает в себя две центриоли и центросферу. Центриоль представляет собой цилиндр, стенка которого образована девятью группами из трех слившихся микротрубочек (9 триплетов), соединенных между собой через определенные интервалы поперечными сшивками. Центриоли объединены в пары, где они расположены под прямым углом друг к другу. Перед делением клетки центриоли расходятся к противоположным полюсам, и возле каждой из них возникает дочерняя центриоль. Они формируют веретено деления, способствующее равномерному распределению генетического материала между дочерними клетками. В клетках высших растений (голосеменные, покрытосеменные) клеточный центр центриолей не имеет. Центриоли относятся к самовоспроизводящимся органоидам цитоплазмы, они возникают в результате дупликации уже имеющихся центриолей. Функции: 1) обеспечение расхождения хромосом к полюсам клетки во время митоза или мейоза, 2) центр организации цитоскелета.
Органоиды движения
Присутствуют не во всех клетках. К органоидам движения относятся реснички (инфузории, эпителий дыхательных путей), жгутики (жгутиконосцы, сперматозоиды), ложноножки (корненожки, лейкоциты), миофибриллы (мышечные клетки) и др.
Жгутики и реснички — органоиды нитевидной формы, представляют собой аксонему, ограниченную мембраной. Аксонема — цилиндрическая структура; стенка цилиндра образована девятью парами микротрубочек, в его центре находятся две одиночные микротрубочки. В основании аксонемы находятся базальные тельца, представленные двумя взаимно перпендикулярными центриолями (каждое базальное тельце состоит из девяти триплетов микротрубочек, в его центре микротрубочек нет). Длина жгутика достигает 150 мкм, реснички в несколько раз короче.
Миофибриллы состоят из актиновых и миозиновых миофиламентов, обеспечивающих сокращение мышечных клеток.
Дата добавления: 2016-05-30; просмотров: 5750;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
