Предмет экологии насекомых 9 глава
Тем не менее диапауза нередко наблюдается и у тропических видов. В этом случае задержка развития может быть довольно кратковременной по сравнению с умеренными широтами. Не ясен вопрос о сезонности у арктических насекомых. Активная жизнь насекомых в Арктике может длиться немногим более одного месяца, но и там наблюдается чередование видов насекомых в течение сезона. Большинство арктических насекомых на протяжении своей жизни зимуют неоднократно, но неизвестно, всегда ли зимующая стадия находится в состоянии диапаузы.
Диапауза может быть эмбриональной, личиночной, пронимфалъной, куколочной или имагинальной. Нет определенной связи между стадией развития, на которой наблюдается диапауза, и систематическим положением насекомого. Так, эмбриональная диапауза часто встречается у саранчовых и некоторых бабочек, в том числе у тутового Bombyx mori L. и японского (Antheraea jamamai) шелкопрядов. Однако у дубового шелкопряда Antheraea pernyi диапаузируют куколки.
Личиночная диапауза типична для многих бабочек (боярышница, златогузка, сосновый шелкопряд), а также для многих мух. Пронимфальная диапауза возможна у бабочек (кукурузный мотылек) и у многих пилильщиков. Имагинальная диапауза типична для многих жуков, комаров, например малярийного, некоторых бабочек, особенно булавоусых. Есть и такие виды, у которых в зависимости от конкретных условий и прежде всего климата диапауза может быть приурочена к разным стадиям развития.
В принципе насекомое диапаузирует именно на той стадии, которая по своему образу жизни или плотности покровов наиболее защищена от внешних воздействий. Так, очень распространена диапауза яиц и куколок, особенно если они находятся в почве или подстилке. Диапауза личинок наблюдается обычно в том случае, если они обитают в почве, или же внутри животных или растительных тканей. Имаго жуков с их жесткими покровами диапаузируют в связи со своей способностью уходить в глубокие укрытия.
У моновольтинных видов диапауза включается как необходимый этап развития, у поливольтинных же диапауза имеет место только у тех генераций, которые попадают на неблагоприятное время года. Правда, многие виды, моновольтинные в северной части своего ареала, становятся поливольтинными в южной.
В случае же длительного развития диапауза неоднократно включается в жизненный цикл. Так, у таракана Ectobius lapponicus L. в первую зиму диапаузирует яйцо, а во вторую – обычно личинка IV возраста (V.K.Brown, 1973). Довольно часто диапауза наблюдается в пределах одного цикла и зимой, и летом. Например, у одного из видов комаров Aedes летом диапаузируют яйца, а зимой личинки (R.G.Jordan, 1980). У некоторых бабочек–парусников летняя диапауза переходит в зимнюю и активное развитие имеет место только весной. Долгоживущие имаго могут диапаузировать многократно на протяжении своей жизни.
Ряд примеров, показывающих разнообразное включение диапаузы в жизненные циклы, приводит в своей монографии В. А.Заславский (1984). Самый простой здесь случай – когда диапауза развивается у поливольтинного вида, в поколении, приходящемся на конец лета или начало осени. Иногда же диапауза наблюдается и зимой, и летом, причем соответственно развитие одного поколения происходит весной, а другого – осенью. У моновольтинных видов диапауза может включаться как обязательный этап или только в зимнее время, или дважды на протяжении цикла развития – зимой и летом.
Индукция диапаузы внешними факторами
Наиболее распространенный случай – факультативная диапауза, возникающая перед наступлением неблагоприятного сезона. Лишь у некоторых моновольтинных видов диапауза становится облигатной и ее наступление не зависит от условий. Устойчивость к неблагоприятным условиям различна на разных стадиях развития, поэтому виду с факультативной диапаузой необходимо подогнать свое развитие к сезону таким образом, чтобы диацаузирующая стадия совпала во времени с неблагоприятными условиями.
Вполне логично было бы предположить, что основным фактором, контролирующим наступление диапаузы, является температура. Действительно, каждый сезон характеризуется своим уровнем температуры, а экспериментальные данные показывают, что есть определенная связь между уровнем температуры и наступлением диапаузы. Заметим, однако, что из года в год сезонный ход температуры несколько различен, а за теплой погодой может последовать резкое похолодание.
Другой возможный фактор, индуцирующий диапаузу, – это качество пищи. Так, мальвовая моль Gelechia malvella Hb. при питании молодыми завязями, цветами и листьями развивается без диапаузы, если же употребляет в пищу зрелые семена растений – всегда диапаузирует. Однако, диапауза имеет место в жизни очень многих насекомых, в том числе и тех, которые не питаются растительной пищей.
В 1920 г. американцы – химик В.Гарнер и физиолог растений Х.Аллард – опубликовали статью, ставшую важнейшим этапом в истории биологии. Они впервые показали, что организм способен реагировать на длину светового дня, изменяя в зависимости от нее характер своего развития. Перед исследователями была поставлена задача, которую мы назвали бы сейчас узко прикладной: почему один перспективный сорт табака, независимо от срока его посева, на севере США зацветает настолько поздно, что наступающие морозы убивают его семена. Исследователи перебрали все возможные варианты экспериментов, среди которых, конечно, температура стояла на первом месте, а длина светового дня – на одном из последних. Однако, оказалось, что если эти растения, выращиваемые при длинном летнем световом дне севера США, затенять, начиная с–16 ч дня, то они рано цветут и дают семена в летнее время, даже если стоит холодная погода. Гарнер и Аллард провели подобные эксперименты и с другими растениями и четко показали, что этапы развития растения определяются длиной светового дня.
Спустя три года вышла работа (S.Marcovitch, 1923), показавшая, что формообразование у тлей также определяется фотопериодом. Позже было обнаружено (M.Kogure, 1933), что эмбриональная диапауза у тутового шелкопряда определяется длиной светового дня, при которой развиваются яйца родительского поколения. В 1941 г. доцент кафедры энтомологии Московского университета Н.С.Андрианова нашла, что куколочная диапауза у китайского дубового шелкопряда Antheraea pernyi Guer. диктуется длиной светового дня, при которой выращиваются гусеницы.
И только, начиная с 50–х гг. нашего века, стало лавинообразно увеличиваться количество работ, описывающих фотопериодическую реакцию самых разнообразных насекомых. Здесь немалую роль сыграла школа, созданная заведующим кафедрой Ленинградского университета А.С.Данилевским (1961). В принципе, такого влияния светового дня на насекомых и следовало ожидать, поскольку длина светового дня зависит от астрономических причин и является самым точным и надежным сигналом, показывающим наступление того или иного сезона.
 Влияние же других факторов обычно накладывается на фотопериодическую реакцию и ее модифицирует. Так, на рис.8 показана зависимость доли диапаузирующих куколок бабочки капустницы от температуры при разной длине светового дня. Можно видеть, что постоянный свет препятствует диапаузе при любой температуре. При 12–часовом световом дне низкая температура стимулирует появление диапаузы, высокая же полностью ее устраняет. В постоянной же темноте (фотопериод равен нулю) кривая почти такая же, как при 12–часовом фотопериоде, но существенно смещенная в сторону более низких температур.
Влияние же других факторов обычно накладывается на фотопериодическую реакцию и ее модифицирует. Так, на рис.8 показана зависимость доли диапаузирующих куколок бабочки капустницы от температуры при разной длине светового дня. Можно видеть, что постоянный свет препятствует диапаузе при любой температуре. При 12–часовом световом дне низкая температура стимулирует появление диапаузы, высокая же полностью ее устраняет. В постоянной же темноте (фотопериод равен нулю) кривая почти такая же, как при 12–часовом фотопериоде, но существенно смещенная в сторону более низких температур.
Рис.8. Зависимость доли куколок белянки Pieris brassicae L. в состоянии диапаузы от температуры и длины светового дня (по А. С.Данилевскому, 1961): 24 ч – постоянное освещение, 12 ч – короткий фотопериод, 0 ч – постоянная темнота
Таким образом, температура – важный фактор в индукции диапаузы, проявляющийся на фоне фотопериодизма. Возможны и случаи, как, например, у наездника–яйцееда Trichogramma pintoi Voegele., когда реакция на температуру более выражена, чем на фотопериод. Особенно важную роль играет температура в становлении диапаузы у таких объектов, которые обитают глубоко в почве или в древесине, куда практически не проникает свет.
Питание также существенно сказывается на становлении диапаузы, модифицируя влияние фотопериода. Так, у златоглазки Chrysopa cornea St. короткий световой день индуцирует диапаузу. Однако даже и при длинном световом дне недостаток тлей, которыми они питаются, приводит к проявлению диапаузы у 60–80% особей (MJ.Tauber et al., 1984). Аналогичным образом недостаток тлей в летнее время индуцирует диапаузу (эстивацию) у божьих коровок. Перед наступлением зстивации, как и перед зимней диапаузой, резко возрастает стремление этих жуков к миграциям (В.П.Семьянов, 1986). Как отмечалось выше, у фитофагов диапауза очень часто связана с изменениями биохимического состава растений в течение сезона (рис.9). В целом изменение пищи или ее недостаток чаще влияют на субтропических и тропических насекомых, где сезонные изменения светового дня не столь значительны.

Большую роль в регуляции диапаузы может играть содержание влаги в пище и в окружающем субстрате. Так, диапауза яиц многих саранчовых продолжается до тех пор, пока не выпадет дождь. Подобная регуляция синхронизует выход личинок с появлением свежей растительности. Для начала эмбриогенеза у комаров Aedes также необходимо увлажнение. Известно, что самки этих комаров откладывают яйца по краям пересыхающих луж. Иногда наличием влаги определяется не диапауза, которая индуцируется фотопериодом, а возможность развития после диапаузы, как имеет место у лугового мотылька Loxostege sticticalis L.
Рис.9. Фотопериодическая реакция хлопковой совки Helicoverpa armigera (Hubner) при питании гусениц разным кормом: / – коробочками хлопчатника, // – листьями хлопчатника. Ось абсцисс – фотопериод в ч, ось ординат – доля куколок в состоянии диапаузы (по Н.И.Горышину, 1958)
Еще один важный фактор, способный сам по себе вызвать диапаузу, – это плотность популяции. Обычно высокая плотность способствует появлению диапаузы. Так, у насекомых – вредителей запасов (Ephestia cautella Wlk., Plodia interpunctella Hb.) скопление личинок, обычно возникающее при недостатке питания, приводит к их диапаузе, а в результате – к длительному сохранению популяции (H.Tsuji, 1963). Интересно, что у жуков–кожеедов Trogoderma variabile Ball, имеет место противоположный эффект. Личинки этого вида могут развиваться только в группах, у одиночных же личинок возникает диапауза (A.Elbert, 1979). Эффекты, связанные с плотностью популяции, а также с влажностью, далеко не всегда совпадают с определенным сезоном. Следовательно, диапауза – приспособление к любым неблагоприятным условиям, а не только к их сезонной смене.
Сигналы, индуцирующие диапаузу, могут быть различными даже у разных географических популяций одного и того же вида. Так, хлопковая моль Pectinophora gossypiella Saund., живущая в северной части своего ареала, реагирует на фотопериод, а в более южных районах – на температуру и качество пищи. Однако слабая реакция на фотопериод сохраняется и у популяций, обитающих около экватора (A.K.Raina and oth., 1981).
Фотопериодическая реакция (ФПР)
Фотопериодическая реакция – это реакция организма на длину светового дня в суточном цикле. Длина светового дня равна интервалу времени между моментами, когда освещенность становится выше пороговой и когда она падает ниже этого порога. Пороговая освещенность очень невелика, около 0,1 лк, что является типично сумеречной освещенностью. Наступление этого уровня строго приурочено к определенному времени и очень мало изменяется из-за облачности. Таким образом, для насекомого длина светового дня почти не зависит от погоды. Очень важно для точного измерения времени также и то, что ФПР, за редкими исключениями, не связана с реальным уровнем освещенности в течение светового дня. Помимо реакции на абсолютную длину светового дня во многих случаях имеет значение также и возрастание или уменьшение длины дня, на чем мы остановимся несколько позже.
Кривые ФПР показывают процент диапаузирующих особей в зависимости от длины фотопериода. Обычно рассматривается альтернатива: диапауза индуцирована – диапауза отсутствует. Оговорим, однако, что во многих случаях фотопериод влияет не только на саму индукцию, но и на длительность диапаузы (златоглазка Chrysopa cornea St., бабочка–медведица Spilarctia imparilis, муха Calliphora vicina R.D.). Чаще всего переход от фотопериодов, индуцирующих диапаузу, к не индуцирующим довольно резок – в пределах одного часа. В отдельных же случаях, как у некоторых малярийных комаров рода Anopheles этот переход является очень постепенным (Е.Б.Виноградова, 1960). Такая генетическая изменчивость ФПР приводит к своеобразному расслоению популяции, т.е. к появлению дополнительных поколений. В случае благоприятной осени эти дополнительные поколения увеличивают численность популяции, в неблагоприятные же годы эти комары погибают, но остаются, как основа, диапаузирующие линии с меньшим числом поколений за год. Итак, ширина переходной зоны кривой ФПР отражает неоднородность в реакциях насекомых одной популяции на фотопериод. Фотопериод, индуцирующий диапаузу у 50% особей называют критической длиной светового дня.
Возможны два типа ФПР: длиннодневная реакция и короткодневная (рис. 10 и 11). В первом случае при длинном световом дне имеет место развитие, а при коротком оно сменяется диапаузой. Длиннодневная ФПР типична для многих поливольтинных видов, имеющих факультативную диапаузу, например, колорадского жука, бабочки–капустницы, щавелевой совки. Этот тип реакции кажется наиболее логичным: сокращение длины светового дня – предупреждение о приближении осенних и зимних холодов. Но диапауза жизненно необходима насекомому и при летнем покое (эстивации). В этом случае именно длинные фотопериоды должны стимулировать ее наступление.
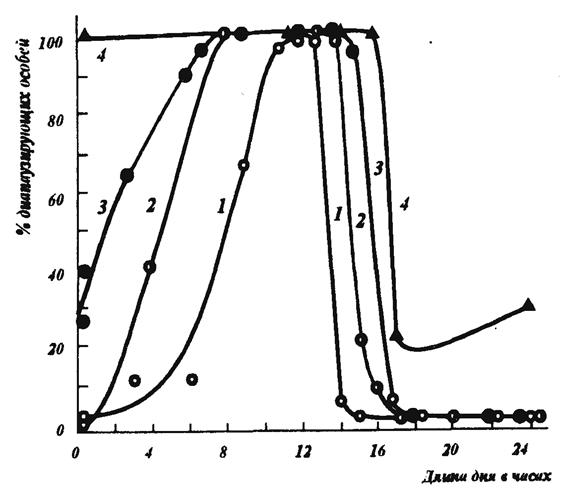
Рис.10. Длиннодневная фотопериодическая реакция насекомых. 1 – листовертка Laspeyre'sia molesta Busck. при 24° С; 2 – белянка Fieris brassicae L. при 24° С; 3 – щавелевая совка Acronycta rumicis L. при 26° С; 4 – колорадский жук Leptinotarsa decemlineata Say. при 24° С (по А.С.Данилевскому, 1961)

Рис.11. Короткодневная фотопериодическая реакция насекомых 1–цикадка Stenocranus minutes F. при 20°С, 2 – тутовый шелкопряд Bombyx тоri L. при 15° С (А.С. Данилевскому, 1961)
Кроме того, реакция на фотопериод совсем не обязательно должна проявляться на следующей стадии и может быть оттянута до следующего поколения. В таких случаях у насекомых наблюдается короткодневный тип реакции, когда короткий световой день стимулирует развитие, а длинный – индуцирует диапаузу. Он более типичен для видов, обитающих к югу от умеренной зоны.
Возможно и сочетание длинно– и короткодневной реакции, так называемый промежуточный тип ФПР, когда развитие может протекать лишь в сравнительно узком диапазоне фотопериодов. Такая ФПР наблюдается, например, у бабочки–желтогузки Euproctis similis Fuessly.
Описанные выше кривые фотопериодической реакции были получены в лаборатории при не меняющихся на протяжении развития насекомого фотопериодах. Искусственность таких опытов очевидна, так как в природе длина светового дня никогда не бывает стабильной, но всегда либо увеличивается, либо уменьшается. Показано, однако, что у многих насекомых эти постепенные изменения фотопериода, пока они не доходят до критической длины, не играют никакой роли в индукции диапаузы (Т.А.Волкович, 1986). Но у ряда насекомых ФПР существенно меняется в зависимости от направления изменений фотопериода. Такую реакцию на изменение длины дня называют ступенчатой. Наиболее распространен короткодневно–длиннодневный тип этой реакции, когда увеличение длины светового дня способствует развитию, а уменьшение – диапаузе. Такая ступенчатая реакция обычно совпадает с длиннодневной реакцией на: константный фотопериод и возможна у активных летом насекомых.
В некоторых случаях реакция на константный фотопериод вообще практически отсутствует. Яркий пример подобной реакции дают божьи коровки Chilocorus bipustulatus L. (В.А.Заславский, 1984). При константных фотопериодах у этих жуков обнаруживается лишь очень слабо выраженная промежуточная реакция (рис.12). Жуки диапаузируют практически при любой длине светового дня. Если же этих жуков поместить в камеру с коротким световым днем (9 ч), а затем переносить в разные тоже константные световые режимы, то окажется, что при фотопериодах меньше 16 ч диапауза будет продолжаться, а при фотопериодах больше 16 ч она будет устранена (рис. 13).
Длиннодневно–короткодневный тип ступенчатой реакции характеризуется активным развитием при уменьшении длины светового дня. Такой тип реакции встречается у насекомых с летней диапаузой и осенним развитием.
У одного и того же насекомого возможно сочетание обоих типов ступенчатых реакций. Так, капустная совка Mamestra brassicae L. в Японии имеет и зимнюю, и летнюю диапаузу. При переносе гусениц из камеры с 12–часовым фотопериодом в 16–часовой возникает летняя куколочная диапауза, а при обратном изменении от 16 ч к 12 имеет место зимняя диапауза (S.Yagi, 1975). В отличие от коровки Chilocorus, у которой при стабильном фотопериоде практически все особи диапаузируют, у японской популяции капустной совки при 14–часовом стабильном фотопериоде развитие протекает без диапаузы.

Рис.12. Фотопериодическая реакция божьей коровки Chilocorus bipustulatus L. на константные фотопериоды. Ось ординат – процент особей в состоянии диапаузы (по В.А.Заславскому, 1984)

Рис.13. Двухступенчатая фотопериодическая реакция божьей коровки Chilocorus bipustulatus L. Ось ординат – процент особей в состоянии диапаузы 1– предварительное нахождение жуков в постоянной темноте, 2 – то же при предварительном фотопериоде 6 ч, 3 – то же при предварительном фотопериоде 9 ч (по В.А.Заславскому, 1984)
Как уже отмечалось выше, ступенчатые реакции могут влиять на наступление диапаузы даже при изменении фотопериода в пределах зон выше или ниже критического порога. Так, у златоглазки Chrysopa cornea St. фотопериоды 14 и 18 ч – выше порога. Однако резкий переход от 18–часового фотопериода к 14–часовому индуцирует диапаузу у 29% особей (M.J.Tauber et al., 1984). Обратная картина наблюдается у луковой мухи Hylemia antiqua Mg. Здесь увеличение фотопериода в зоне ниже критической снижает процент диапаузирующих особей, укорочение же фотопериода в зоне выше критической не влияет на уровень диапаузы (Е.Б Виноградова, 1978).
Стадия развития, чувствительная к фотопериоду
Чувствительность к фотопериоду чаще всего проявляется на стадии, предшествующей диапаузе. Так, диапауза на стадии куколки обычно индуцируется в зависимости от режима, в котором были выращены гусеницы. Диапауза у имаго часто определяется фотопериодическими условиями, в которых находилась куколка, но иногда условиями содержания личинок. Чувствительность к фотопериоду может долго сохраняться у имаго насекомых, живущих несколько лет, например у жужелиц, и диапауза у них может повторяться многократно.
Сложнее вопрос об, индукции эмбриональной диапаузы. У тутового шелкопряда, например, яйца диапаузируют в том случае, если яйца и гусеницы первых возрастов родительского поколения развивались при длинном световом дне. У тлей диапауза яиц может определяться теми фотопериодическими условиями, в которых находилось поколение "бабушек".
В тех случаях, когда фотопериодические и температурные воздействия не отражаются на развитии самой особи, но изменяют реакцию дочернего поколения, такой эффект называют материнским влиянием. При этом возможны все варианты: от жесткой зависимости от условий, в которых содержалось родительское поколение, как у тутового шелкопряда, до сравнительно небольшой модификации ФПР дочернего поколения.
Материнское влияние возможно и при температурном управлении диапаузой. Так, у трихограммы Trichogramma pintoi Voegele, чем выше температура, при которой развивалось родительское поколение, тем ниже склонность к диапаузе у дочернего. У Trichogramma evanescens Westw., наоборот, высокая температура способствует появлению диапаузы в дочернем поколении.
Конечно, не всегда можно отличить материнское влияние от воздействия фактора на развивающийся эмбрион яйца, которое еще находится в теле матери. Если у мясных мух Calliphoridae имеет место материнское влияние (Т.П.Богданова и др., 1978; Е.Б.Виноградова, 1984,1991), то у мух семейства Sarcophagidaе удается показать восприятие фотопериодической информации сквозь покровы тела матери непосредственно самим эмбрионом (Е.Б.Виноградова, 1978).
Продолжительность времени, в течение которого организм насекомого чувствителен к факторам, индуцирующим диапаузу, различна у разных насекомых и меняется от нескольких дней в течение жизненного цикла (комары рода Aedes) до всего цикла развития (муравьиный лев Myrmeleon formicarius L.). В ряде случаев чувствительность к фотопериоду сохраняется и после наступления диапаузы и увеличивает ее длительность. Так, чувствительность гусениц Ostrinia nubilalis Hb. к фотопериоду исчезает только к середине декабря, т.е. к тому времени, когда наступает устойчивое похолодание. Известны также факты длительного поддержания летней диапаузы длинным световым днем. Подробнее на вопросах о поддержании и прекращении диапаузы мы остановимся в следующих разделах.
Для получения определенного фотопериодического эффекта совсем не обязательно, чтобы насекомое воспринимало один и тот же фотопериод в течение всего интервала времени, когда оно к нему чувствительно. Так, для индукции диапаузы у капустной совки Mamestra brassicae L. достаточно воспринять всего 14–15 коротких фотопериодов (критический пакет фотопериодической информации – В.П.Тищенко, 1977). Остальные же фотопериоды могут быть как угодно длинными. Однако, например, у щавелевой совки Acronycta rumicis L. наступление диапаузы определяется конечным режимом, в котором находились гусеницы этого вида, чувствительные к фотопериоду.
Фотопериодическая реакция и температура
В предыдущих разделах мы уже останавливались на некоторых примерах взаимодействия ФПР и температуры. У большинства насекомых с длиннодневным типом фотопериодической реакции повышение температуры приводит к уменьшению критического фотопериода и устранению диапаузы (рис. 14). На каждые 5° критический фотопериод сдвигается на 1–1,5 ч, что соответствует естественным изменениям фотопериода за 2–4 недели. В результате увеличивается срок, в течение которого возможно развитие насекомого. При температурах, близких к сублетальным, развитие идет практически всегда без диапаузы. Такая зависимая от температуры лабильность ФПР дает возможность некоторым видам проникать из умеренной зоны в субтропическую, где фотопериод всегда непродолжителен. Наоборот, "усиление" ФПР при низкой температуре способствует сохранению вида при необычно раннем наступлении осени.
У видов с короткодневной ФПР, как и следует ожидать, реакция на температуру противоположна и высокая температура способствует наступлению диапаузы.
 Возможны и другие варианты воздействия температуры на ФПР. Так, у восточной плодожорки Grapholitha molesta Busck. и хлопковой совки Helicoverpa armigera (Hubner) диапауза может быть вызвана коротким фотопериодом только в диапазоне температур 20–25°. Такая связь индукции диапаузы с довольно высокой температурой скорее всего объясняется южным происхождением этих видов. Наоборот, у бабочки–медведицы Spi–losoma menthastri Esp. развитие без диапаузы, зависящее также от ФПР, возможно только в пределах температур 20–27°.
Возможны и другие варианты воздействия температуры на ФПР. Так, у восточной плодожорки Grapholitha molesta Busck. и хлопковой совки Helicoverpa armigera (Hubner) диапауза может быть вызвана коротким фотопериодом только в диапазоне температур 20–25°. Такая связь индукции диапаузы с довольно высокой температурой скорее всего объясняется южным происхождением этих видов. Наоборот, у бабочки–медведицы Spi–losoma menthastri Esp. развитие без диапаузы, зависящее также от ФПР, возможно только в пределах температур 20–27°.
Рис.14. Зависимость критической длины светового дня у щавелевой совки Acronycta rumicis L. от температуры, при которой происходило развитие гусениц (по А.С.Данилевскому, 1961)
Итак, ФПР обычно имеет место в диапазоне температур, оптимальных для роста и развития. В некоторых случаях, однако, ФПР представляется совсем не связанной с уровнем температуры. С другой стороны, известен ряд видов, у которых наступление диапаузы определяется прежде всего температурой. Подобное влияние температуры четко выражено у таких объектов, как, например, живущие в почве личинки некоторых пластинчатоусых жуков, у находящихся в почве яиц саранчовых. К числу таких насекомых относятся также обитающие в древесине или под корой личинки жуков, некоторые вредители запасов, например гусеницы бабочки Plodia interpunctella Hb., a также ряд тропических видов насекомых.
Как и в случае с фотопериодом, на развитие может влиять не только сам уровень температуры, но и направление ее изменений. Так, повышение температуры уменьшает процент диапаузирующих личинок мух Sarcophaga и Calliphora. Понижение температуры в сочетании с уменьшением длины светового дня резко увеличивает склонность куколок бабочки Heliothis zea к диапаузе.
Суточный ход температуры может также влиять на сезонное развитие. Так, стеблевой кукурузный мотылек Ostrinia nubilalis Hb. на 100% диапаузирует при коротком фотопериоде и постоянной температуре, а также при температурном цикле 31° днем и 21° ночью. Если же цикл температуры инвертирован (31° ночью и 21° днем), то доля диапаузирующих особей резко падает, при любом фотопериоде (S.Beck, 1962). Следовательно, индукция диапаузы зависит от уровня температуры в ночное время.
При содержании насекомых в постоянной темноте продолжительность теплого периода в течение суток может играть роль, аналогичную фотопериоду. Здесь кривая индукции диапаузы в зависимости от продолжительности термопериода очень сходна с кривой ФПР.
Географическая изменчивость ФПР
У видов, имеющих большой ареал, фотопериодическая реакция различна в разных его частях. На рис.15 показаны результаты изучения ФПР у разных географических популяций широко распространенной совки Acronycta rumicis L. (А.С.Данилевский, 1961). Сравнивали ФПР популяций из Сухуми (43° с.ш.), Белгорода (51° с.ш.), Витебска (55° с.ш.) и Петербурга (60° с.ш.). Чем севернее происхождение популяции, тем больше критическая длина светового дня. Здесь очевидно четкое соответствие индукции диапаузы реальной длине светового дня на данной широте в конце лета и начале осени. У самой северной петербургской популяции значительная часть особей диапаузирует и в условиях длинного дня, т.е. эта популяция имеет четкую тенденцию к моновольтинизму и формированию облигатной диапаузы.
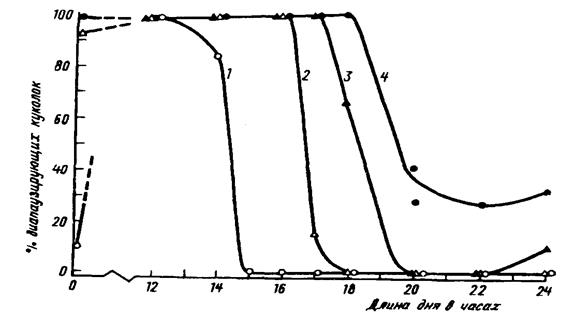
Рис.15. Географическая изменчивость фотопериодической реакции щавелевой совки Acronycta rumicis L. 1 – сухумская, 2 – белгородская, 3 – витебская и 4 – петербургская популяции (по А.С.Данилевскому, 1961)
При воспитании этих гусениц в открытых вольерах на широте Петербурга выявилось следующее. Все куколки петербургской популяции диапаузируют, начиная с конца июля. Диапаузирующие гусеницы белгородской популяции появляются в сентябре, а сухумской – только в октябре. Следовательно, потомство южных по происхождению бабочек, случайно попавших в более северные районы, будет запаздывать с наступлением диапаузы и, скорее всего, погибнет. Бабочки же северных популяций, попавшие на юг с его коротким днем, должны будут диапаузировать практически круглогодично и не смогут развиваться. Кроме того, длительная диапауза при высокой температуре всегда приводит к гибели от высыхания и истощения жировых запасов.
Подобная же изменчивость ФПР проявляется при сравнении популяций, обитающих на одной географической широте, но в районах с разным климатом. Владивосток и Сухуми расположены на одной широте, однако климат Владивостока значительно суровее. Соответственно, у белянки Pieris rapae L. в Сухуми критическая длина светового дня равна 12 ч, а во Владивостоке – 15ч, т.е. в более суровом климате диапауза наступает значительно раньше (А.С.Данилевский, 1961).
Можно было ожидать, что сходные закономерности должны выявляться при анализе ФПР популяций одного вида, обитающих на разной высоте над уровнем моря. Однако, если у кавказского подвида белянки Pieris rapae meridionalis, живущего в долинах у морского побережья, имеет место четкая длиннодневная ФПР, то у горного подвида Pieris rapae bryoniae имеется тенденция к моновольтинизму, ФПР почти не выражена, а при 18° диапаузируют все куколки независимо от фотопериода (А.С.Данилевский, И.А.Кузнецова, 1968).
Таким образом, географическая изменчивость ФПР значительно затрудняет переселения вида внутри ареала. Особенности ФПР необходимо учитывать при попытках интродукции и акклиматизации новых для фауны видов, специально подбирая при этом определенные исходные популяции. Кроме того, особенности ФПР существенно тормозят распространение карантинных видов.
Реактивация
Переход от диапаузы к активному развитию называют реактивацией. Реактивация, так же как и начало диапаузы, должна быть приурочена к строго определенному сезону. Например, зимняя диапауза у большинства видов умеренной зоны индуцируется коротким световым днем и холодом. Она начинается в конце лета или осенью. Очевидно, что развитие таких насекомых может возобновиться не раньше, чем весной, с наступлением устойчивого потепления. Температура в укрытиях в зимнее время в умеренной зоне всегда ниже порога развития, и зимних оттепелей недостаточно, чтобы провоцировать несвоевременное развитие насекомых и, соответственно, их выход из состояния зимовки. Однако если насекомых уже в начале декабря перенести в тепло, у многих из них возобновляется нормальное развитие (например, щавелевая к хлопковая совки). У ряда видов такая реактивация все же возможна не раньше чем весной или даже в начале лета.
Дата добавления: 2016-06-29; просмотров: 1613;
Поиск по сайту
Узнать еще
- I.1.1 ПРЕДМЕТ МОЛЕКУЛЯРНОЙ ФИЗИКИ
- Активные – воздействуют на предмет труда, перемещают его, контролируют ход производства (машины, оборудование, т/с, инструмент)
- Актуальность изучения экологии
- Алфавитно-предметный указатель
- Аналитическая химия и химический анализ. Предмет и задачи аналитической химии. Классификация методов химического анализа.
- Аналогично работают ПТК СВШД и системы обнаружения случайных предметов (СОСП).
- Анатомо-физиологические аспекты потребностей человека. Задачи изучаемого предмета.
- Антропогенное влияние на природу. Проблема экологии.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
