Предмет экологии насекомых 8 глава
Другой важный приспособительный момент – синхронизация вылета насекомых разного пола. Это особенно четко проявляется на примере поведения афагов, таких как поденки и бабочки–тонкопряды. Очевидно, что синхронный выход позволяет этим насекомым встретиться и оставить потомство с наименьшей потерей особей. Лет некоторых видов поденок продолжается на протяжении всего 20–30 мин. Интересно, что вылет поденок, размножающихся партеногенетически, значительно более растянут во времени, чем тех видов, у которых известны и самцы, и самки. Ту же роль играет эндогенное управление ритмом выделения полового феромона у самок бабочек.
Эндогенный ритм имеет большое значение и в пространственной ориентации насекомых. О способности насекомых вносить регулярную временную поправку на движение небесного ориентира по небосводу – астротаксисе – говорилось выше. Эндогенный ритм здесь как бы переключает в определенном порядке омматидии, фиксирующие небесный ориентир, например Солнце.
Помимо пространственной ориентации, пчелы используют эндогенный ритм, приспосабливаясь ко времени выделения нектара разными цветками. Пчелы способны запомнить время, когда они могут получить корм, и прилетают за ним в определенное время суток. Такое поведение называют рефлексом на время.
Наконец, эндогенный ритм имеет большое значение в сезонном фотопериодизме, о котором речь пойдет в следующих разделах. Один из возможных способов определения длины фотопериода основан на сопоставлении эндогенного ритмического процесса со световым циклом (В.П.Тыщенко, 1977).
Суточный ритм чувствительности организма насекомого
Эндогенный суточный ритм отражается на всех физиологических функциях организма насекомого. Это особенно четко проявляется на примере суточных изменений чувствительности рецепторов и способности всего организма в целом реагировать на изменения условий.
Поместив насекомых в постоянную темноту и наблюдая их реакцию на включение света в разное время суток, легко убедиться, что и скорость реакции и число реагирующих особей меняются. Так, для многих дневных насекомых в дневное время свет является резко возбуждающим фактором, но к вечеру уровень реакции снижается и остается на низком уровне до наступления утра. Эти изменения реакции на свет, несомненно, связаны с суточным ритмом чувствительности глаза насекомого. Форма электроретинограммы глаза и пороговая интенсивность света меняются в течение суток даже в постоянных условиях лаборатории. У ночных насекомых чувствительность глаза резко возрастает к вечеру, у дневных ритм чувствительности более разнообразен (Г.А.Мазохин–Поршняков, 1965). Описаны также суточные изменения работы обонятельных рецепторов насекомых, особенно четко проявляющиеся в реакции самцов на феромон самок.
Помимо чувствительности глаза в течение суток меняется большинство физиологических и биохимических характеристик организма, поэтому чувствительность к любым воздействиям оказывается, как правило, неодинаковой в разное время суток. Чувствительность насекомого к экстремальным температурам, как высоким, так и низким, максимальна во время активности насекомого, независимо от образа его жизни. Аналогичен ритм чувствительности насекомого к рентгеновским лучам. Так, у дрозофил смертность максимальна при облучении во второй половине дня, а у яблоневой плодожорки – вечером, т.е. в то время, когда эти насекомые наиболее активны (В.П.Приставко, Н.Э.Семьянова, 1973).
На протяжении суток меняется и чувствительность насекомых к ядохимикатам. Иногда эти изменения исключительно резки. Некоторые долгоносики сразу же после включения, света утром мало чувствительны к метилпаратиону и от определенной дозы этого яда погибает не более 10% жуков. Однако спустя три часа та же доза приводит к гибели 90% жуков. Многочисленные исследования показывают, что чаще всего повышенная чувствительность к пестициду имеет место во время максимальной активности насекомого. Все же механизмы воздействия разных пестицидов различны, и время максимальной чувствительности к ним может не совпадать, а в некоторых случаях быть почти в противофазе. Совершенно очевидно, что испытание и применение пестицидов, включая аналоги гормонов и, возможно, вирусные и другие биопрепараты, должны быть приурочены к определенному времени суток. Это время суток не может быть стандартным для каждого пестицида и должно изменяться в зависимости от сезона, вида и географической популяции насекомого. Знание времени наибольшей чувствительности к пестицидам и другим воздействиям позволило бы более рационально организовать борьбу с вредными насекомыми, сократив при этом загрязнение среды.
Факторы среды – датчики времени
Эндогенный ритм не является, однако, чем-то раз навсегда данным организму. Он может подстраиваться к изменяющимся условиям среды за счет сдвига его фазы. Факторы, способные сдвигать фазу эндогенного ритма, называют датчиками времени. Второе обязательное свойство любого датчика времени – строго определенное и постоянное соотношение между его циклом и ритмом животного (постоянный фазовый угол).
Очевидно, что фактор среды, суточный ход которого неотчетлив и постоянно нарушается, не может служить хорошим сигналом для подстройки ритма, т.е. быть датчиком времени. Наоборот, чем точнее повторяется от суток к суткам изменение фактора, тем большую роль может играть этот фактор в синхронизации ритма организма с ритмом среды. Нерегулярно меняющийся фактор может быть только помехой для настройки ритма, поэтому биологические часы (эндогенный ритм) должны быть максимально изолированы от таких факторов. Аналогичным образом эндогенный ритм изолирован и от общих обменных процессов в организме, ход которых неустойчив и может зависеть от многих причин.
Попробуем оценить роль различных факторов среды как датчиков времени.
Свет – наиболее четко и резко меняющийся в течение суток фактор. Ежесуточно освещенность в средних и низких широтах меняется в пределах от десятых долей люкса (в самую яркую лунную ночь) до тысяч люксов (в наиболее пасмурный день). В полярных широтах суточный ход освещенности выражен гораздо слабее (круглосуточно незаходящее солнце). Соответственно и ритм иногда теряет в таких условиях временные ориентиры и в середине лета у некоторых животных регистрируется уже не суточный ритм, а циркадианный, расходящийся по фазе с местным временем. Подобное же явление можно наблюдать и в термостате с постоянным освещением или постоянной темнотой.
В умеренной, а тем более в тропической зоне по уровню освещенности можно довольно точно определять время. Облачность может сдвигать ход освещенности в утреннее и вечернее время не более чем на 10–20 мин. Не случайно, что именно свет является главным фактором, синхронизующим и настраивающим ритм. От случайных же изменений освещенности, например резкого падения освещенности в середине дня от плотной облачности, организм защищен ритмом чувствительности к свету, о котором шла речь в предыдущем разделе. Поэтому настройка ритма может осуществляться как полным световым циклом, так и двумя относительно непродолжительными световыми импульсами, один из которых соответствует началу дня, а другой – его концу (скелетный фотопериод). Для синхронизации ритма отдельных особей также нет необходимости в восприятии полного суточного светового цикла. Так, если куколки дрозофил находятся в полной темноте и в этих же условиях происходило развитие личинок, мухи из этих куколок появляются в любое время суток. Однако достаточно однократной и очень кратковременной засветки культуры, чтобы дрозофилы стали выходить синхронно в определенное время суток, соответствующее тому времени, когда они, еще будучи личинками или куколками, восприняли свет.
Другим важным датчиком времени может быть температура. Этот фактор, как правило, также четко меняется в течение суток, хотя иногда и возможны резкие нарушения его суточного хода. В отличие от освещенности, температура изо дня в день колеблется на разных уровнях. Температура может быть датчиком времени в основном для пойкилотермных животных, в том числе и для насекомых. Здесь также имеют место изменения чувствительности к колебаниям температуры в течение суток, защищающие ритм от помех.
Температура является важнейшим датчиком времени для многих, часто для нелетающих насекомых. Однако в ряде случаев, как, например, в ритме активности комнатной мухи (В.А.Зотов, С.А.Федосов, 1983) или в ритме выхода наездника–трихограммы из куколки (A.Dahyia et al. 1993), оба фактора – и свет, и температура – являются равноценными и взаимно заменяющими друг друга датчиками времени. При этом повышение температуры воспринимается так же, как увеличение уровня освещенности.
Влажность, хотя и является важнейшим фактором в жизни насекомых, обычно не регулирует эндогенные ритмы. Исключение – ритм выхода из яиц гусениц американской белой бабочки, который четко связан с повышением уровня влажности (С. А.Федосов, 1990).
Предполагалось, что некоторые насекомые способны ориентироваться во времени по суточным изменениям гравитации (F.Scneider, 1964). Несомненно, что этот фактор должен обладать четкой астрономической периодичностью. Однако поскольку гравитационное воздействие Луны больше, чем Солнца, то основная информация, которую могли бы воспринимать насекомые, касалась бы не солнечных, а лунных суток (24,8 ч). Именно такая периодичность была обнаружена в пещерах у обитающих там кузнечиков (R.Simon, 1973).
Спокойное геомагнитное поле имеет также довольно регулярные суточные вариации. В годы повышенной солнечной активности возмущения геомагнитного поля наблюдаются почти ежедневно и их амплитуда существенно больше суточных вариаций. Выше мы уже отмечали, что у жуков–кожеедов были отмечены нарушения суточного ритма подвижности, совпадающие с геомагнитными бурями. Скорее всего, здесь наблюдалось только возбуждающее действие геомагнитной бури на насекомое, но не изменения эндогенного ритма. Если бы насекомые ориентировались во времени именно по суточным вариациям геомагнитного поля, их ритм нарушался бы гораздо чаще, чем это действительно имеет место.
Четким суточным ходом обладают также электромагнитные колебания, возникающие в атмосфере, – атмосферики. Тем не менее в наших опытах экранирующие камеры, препятствующие проникновению атмосфериков, не снижали синхронность ритма выхода дрозофил из куколок, а, наоборот, ее повышали.
Итак, по информационной ценности для эндогенного суточного ритма на первом месте оказывается суточный ход освещенности, на втором – температуры, на третьем – влажности воздуха. Участие прочих факторов в регуляции эндогенного ритма сомнительно хотя бы потому, что их регулярный ход часто и существенно нарушается и они оказываются ненадежными источниками информации о времени.
Время потенциальной готовности
Ритм можно представить как чередование активности и покоя. Эндогенный ритм создает готовность к переходу от одного состояния к другому, которая постепенно нарастает. Одновременно резко увеличивается чувствительность к сигнальному фактору, стимулирующему ожидаемое изменение активности. Это время повышенной чувствительности к сигнальному фактору (датчику времени) называют временем потенциальной готовности (ВПГ).
ВПГ начинается с того момента, когда сигнальный фактор способен вызвать переход от покоя к активности (или обратно), а кончается при настолько высокой готовности, что этот переход происходит спонтанно, без внешнего стимула. В природе сигналом служит определенный уровень сигнального фактора, например освещенности. Чем ближе фактор к этому уровню, тем быстрее наступает реакция (переход от покоя к активности или обратно).
Приведем пример, показывающий наличие ВПГ в ритмике. Пустынные жуки–чернотелки Trigonoscelis gigas Reitt. активны дважды в сутки: утром и вечером. Середину дня и ночь они проводят под поверхностью песка. Следовательно, в их ритме имеются два периода покоя (дневной и ночной) и 4 ВПГ (при переходе к активности утром и ее прекращении перед наступлением жары, при переходе к активности вечером и при уходе в песок с наступлением темноты). Для определения положения во времени и длительности интервалов ВПГ, а также сигнальных факторов, управляющих изменениями активности, была предложена следующая методика (В. А.3отов, 1980).
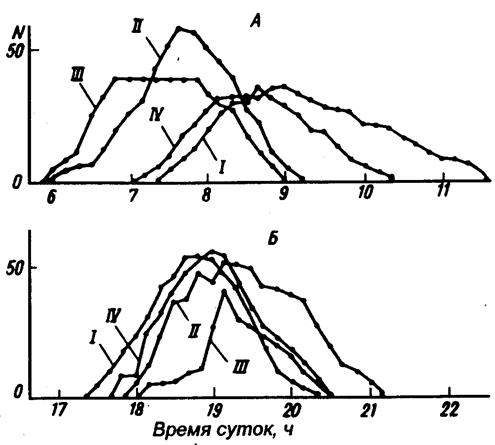
Рис.7. Число жуков Trigonoscelis gigas Reitt. на поверхности почвы в загонах под открытым небом в зависимости от времени суток (по В.А.Зотову, 1980) А – утренний максимум активности, Б – вечерний максимум активности; IV – контроль, дополнительные воздействия: I – затенение, II – освещение, III – нагрев
На первом этапе исследуют ритм в природных условиях: определяют положение максимумов активности во времени и приблизительно намечают положение ВПГ. На втором этапе в искусственных загонах или в садках создают с помощью ламп, нагревателей или затемнения преждевременное или запаздывающее наступление условий, при которых начинается или кончается активность насекомых в природе.
Результаты такого эксперимента, проводившегося под открытым небом с жуками–чернотелками, показаны на рис.7. Можно видеть, что позже всего утром вышли на поверхность песка жуки, не подвергавшиеся никаким дополнительным воздействиям (контроль). Чуть раньше появились жуки в варианте с затенением поверхности песка (под затеняющей поверхность песка черной материей нагрев сначала был даже несколько больше, чем в контроле). Зато почти на полтора часа раньше начиналась активность жуков в вариантах с подогревом песка и дополнительным освещением. Подчеркнем, что осветительные лампы тоже заметно нагревали песок, хотя и в меньшей степени, чем обогреватель.
Следовательно, в течение первого ВПГ основным сигналом является температура. Свет здесь не играет большой роли, так как затенение не задерживает выход жуков по сравнению с контролем.
Рассмотрим, как развертывается поведение жуков в течение второго ВПГ. Раньше всего ушли в песок жуки в вариантах "нагрев" и "свет", позже всего – в варианте "затенение", где нарастание температуры в дальнейшем отставало по сравнению с открытой поверхностью песка в контроле. Следовательно, сигналом в течение ВПГ2 служит скорее всего температура (может быть, и свет). Аналогичные рассуждения приводят к выводу, что сигналом в течение ВПГЗ служит снижение температуры, однако ВПГЗ короче, чем ВПГ1 в течение утреннего пика активности, и различия во времени начала активности здесь не более, чем 40 мин. Во время заката солнца активность прекращается во всех вариантах, за исключением варианта "освещение". Поэтому есть основания предположить, что основной сигнальный фактор в течение ВПГ4 – свет.
Сигнал, поданный преждевременно, не приводит к изменению активности, пока не наступит ВПГ. Так, в тех же опытах нагрев и свет включали в 3 ч ночи. Выход же жуков наблюдался примерно в то же время, как и в предыдущих опытах, т.е. около 5 ч 30 мин.
Может возникнуть сомнение, не связано ли отсутствие реакции во время покоя с их уходом глубоко в песок. Но в опытах, поставленных по аналогичной схеме с комнатными мухами, практически не использующими во время покоя укрытия, были получены очень похожие результаты (В.А.Зотов, С.А.Федосов, 1983). Правда, продолжительность ВПГ у мух гораздо больше, чем у чернотелок, поэтому ритм их активности гораздо лабильнее. В принципе, сумма продолжительностей ВПГ в течение суточного цикла может быть мерой эндогенности ритма – чем больше эта сумма, тем большую роль играют в ритме экзогенные воздействия, т.е. внешние факторы.
Рассматривая суточные ритмы самых различных насекомых, легко обнаружить, что реакция на сигнальный фактор у них также ограничена определенным временем суток. Чаще всего таким сигналом является определенный уровень освещенности, но иногда имеет значение и направление ее изменения. Так, в лаборатории можно вызвать роение комаров-дергунов утром только при повышении освещенности, а вечером – только при ее понижении. В другое время суток вызвать роение оказывается невозможным (J.Syrjamaki, 1968).
В начале интервала ВПГ сигнал вызывает относительно слабую и растянутую во времени реакцию. Ближе к концу ВПГ готовность высока и реакция отличается высоким уровнем и резкостью. ВПГ, очевидно, имеет большое экологическое значение. С одной стороны, благодаря нему организм сопротивляется случайному изменению условий, провоцирующему несвоевременную активность. С другой стороны, ВПГ позволяет организму подстроить ритм к конкретным условиям текущего дня (погоде, сезону)
Циркадианные ритмы
Если насекомое находится в термостате в постоянной темноте или при слабом постоянном освещении, его активность часто остается ритмичной как и при естественных условиях. Однако период такого ритма обычно несколько отклоняется от 24 ч. Такие проявляющиеся в постоянных условиях ритмы, период которых меняется в диапазоне примерно от 22 до 27 ч, называют циркадианными (околосуточными). Период циркадианного ритма индивидуален, но, как правило, закономерно изменяется в зависимости от уровня постоянной освещенности в опыте.
Попытаемся представить, как должны меняться параметры ритма в постоянных условиях, исходя из того, что сдвиги фазы ритма возможны только в течение ВПГ (В.Б.Чернышев, 1984).
Период ритма τ делится на два интервала: время активности α и время покоя р, т.е. α + ρ = τ . В конце каждого из этих интервалов времени возникает готовность к переходу в альтернативное состояние. Назовем интервал готовности в конце времени покоя – ВПГ1 и в конце времени активности – ВПГ2. Сигналы, стимулирующие начало активности и начало покоя, противоположна (так, для дневного насекомого стимул перехода к активности – свет, к покою – темнота).
Примем как аксиому, что интервалы времени от реального начала активности до наступления готовности к покою (только готовности, но не обязательно самого перехода!), а также от реального начала покоя до готовности к активности – постоянные величины. Назовем первый интервал времени – временем обязательной активности – BOA, a второй – временем обязательного покоя – ВОП. Очевидно, что активность может продлеваться за счет запаздывания перехода к покою, т.е. за счет ВПГ2, а покой – за счет прибавления к ВОП времени ВПГ1. Таким образом, если α и р – реально наблюдаемые продолжительности активности и покоя, то:
ΒΟΑ < α < ΒΟΑ + ВПГ2
ВОП < ρ < ВОП + ВПГ1 .
Соответственно максимальный и минимальный периоды ритма, которые возможны на основе этой схемы, следующие:
τ 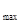 = BOA + ВОП + ВПГ1 + ВПГ2 ,
= BOA + ВОП + ВПГ1 + ВПГ2 ,
τ 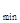 = ΒΟΑ + ВОП.
= ΒΟΑ + ВОП.
Если эта схема верна, то ритм не имеет какого-либо определенного собственного периода, а есть только строго определенный диапазон, в котором этот период может меняться в зависимости от условий. В таком случае ритм всегда является продуктом взаимодействия внутренних эндогенных процессов со средой и всегда включает в себя элемент реакции на среду, даже если она не меняется во времени. Эндогенный ритм без экзогенного компонента невозможен, как невозможно существование животного вне какой бы то ни было среды.
Представим себе, как должен меняться ритм в постоянных условиях в зависимости от уровня освещенности. Рассмотрим только контрастные ситуации: свет и темнота.
Если насекомое ведет дневной образ жизни, то свет будет способствовать его активности, а темнота – покою, т.е. свет будет стимулировать переход от покоя к активности в течение ВПГ1, а темнота – переход к покою в течение ВПГ2. Тогда при постоянном освещении после окончания времени обязательного покоя сразу же с наступлением ВПГ1 начнется активность. В конце же активности свет будет тормозить переход к покою вплоть до окончания ВПГ2. В итоге период ритма дневного насекомого в постоянном освещении равен:
τ 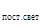 = ВОП + ВОА + ВПГ2.
= ВОП + ВОА + ВПГ2.
Рассуждая подобным же образом, приходим к выводу, что период ритма в постоянной темноте равен:
τ 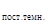 = ВОП + ВПГ1 + ВОА.
= ВОП + ВПГ1 + ВОА.
Если продолжительность ВПГ1 и ВПГ2 одинакова, то период циркадианного ритма не будет зависеть от уровня освещенности в опыте, хотя и не обязательно будет равен 24 ч. Обычно же эти ВПГ различны, соответственно период циркадианного ритма зависит от уровня постоянной освещенности. Чаще ВПГ1 больше ВПГ2, поэтому у дневных насекомых с повышением уровня постоянной освещенности в опыте период циркадианного ритма обычно укорачивается.
Для ночного насекомого будут справедливы те же рассуждения, но так как для его активности более благоприятна темнота, необходимо время активности и время покоя поменять местами. Сигналом, регулирующим ритм, может, конечно, служить не только свет, но и температура.
При рассмотрении изменений ритма реальных насекомых в постоянных условиях картина может усложняться за счет взаимодействия пиков активности в течение суток. Так, у упомянутых выше жуков–чернотелок имеются два пика активности – утренний и вечерний (В.А.Зотов, 1983), которые влияют друг на друга.
4. СЕЗОННЫЕ РИТМЫ
Согласование жизнедеятельности насекомых с сезоном
В умеренной зоне в зимнее время активность насекомых, за редкими исключениями, оказывается невозможной из-за низких температур, опасности замерзнуть. Кроме того, зимой большинство насекомых–фитофагов и хищников не может найти себе пищу. Но и в тропических и субтропических районах сезонность может быть тоже резко выражена. Здесь неблагоприятно для насекомых сухое и жаркое время года.
Сезонность в жизни насекомых часто связана с жизнью растений. Например, появление многих листогрызущих гусениц и тлей четко синхронизовано с весенним распусканием листьев. Если эти насекомые появятся раньше, они погибнут от голода, если позже, – они не смогут воспользоваться молодой и наиболее питательной листвой. Очень четко сезонность выражена у насекомых, питающихся растениями–эфемерами или на цветах в засушливой зоне, где цветение очень непродолжительно. Помимо этого, сезонность может иметь большое значение для репродуктивной изоляции близких видов и снижать межвидовую конкуренцию.
Насекомые приспосабливаются к сезонным изменениям среды разными способами. Во-первых, они могут на неблагоприятное время мигрировать в другую климатическую зону. Во-вторых, они могут зарыться глубоко в почву, где сезонные изменения условий почти отсутствуют. К зимовке или летней засухе часто приспособлена именно та стадия, которая проходит в почве (например, кубышки с яйцами саранчовых, куколки и предкуколки бабочек и других насекомых). В-третьих, насекомые могут выработать повышенную устойчивость к неблагоприятным воздействиям, прежде всего к низким температурам. Эта устойчивость может быть либо постоянной, либо приуроченной к определенной стадии развития и определенному сезону.
Наиболее наглядный способ изложения фенологии того или иного вида – это составление фонограмм (Б.В.Добровольский, 1969), в которых по графам, соответствующим декадам сезона, с помощью условных значков отображается наличие тех или иных стадий развития данного насекомого.
Сезонные миграции насекомых
Известно сравнительно немного примеров дальних миграций насекомых, связанных с сезоном. Дело в том, что насекомые мигрируют в основном с воздушными потоками, т.е. ветром. Миграции способствуют расселению насекомых, но редко бывают направленными. Возможность противостоять ветру и лететь в определенном направлении может быть только у крупных и хорошо летающих насекомых. Мелкие же и плохо летающие могут перемещаться в нужном направлении только небольшими перелетами и непосредственно над покровом растительности, где ветер слабее. Такое перемещение относительно мало эффективно и не дает им возможности мигрировать на большие расстояния.
Увеличение миграционной активности часто связано с недостатком пищи, а также витамина "Е" в питании, который необходим для созревания половых продуктов. Отметим, что даже при случайном направлении перелетов такая миграция, если она достаточно продолжительна, может обеспечить выживание вида с последовательным ежегодным заселением новыми поколениями тех районов, где переживание в неблагоприятный сезон невозможно.
Существенно облегчает миграцию наличие зрительных ориентиров, например темной полосы края леса или очертаний гор. Так, относительно небольшие сезонные перелеты совершают некоторые божьи коровки, которые откочевывают осенью в горные или возвышенные участки местности, где иногда образуют большие скопления. Даже высотное здание Московского университета, по-видимому, может служить центром притяжения для таких миграций. В Калифорнии коровки Hippodamia convergens, обитающие весной в долинах, где в это время много корма, с июня улетают в каньоны Сьерра–Невады, где прохладнее, сохраняется летом много растительности и можно найти тлей (K.S.Hagen, 1962).
Наиболее поразительны сезонные тысячекилометровые миграции бабочки–монарха Danaus plexippus L. Летом этих бабочек можно найти на американском континенте вплоть до Южной Канады, где они успешно размножаются, давая за лето ряд поколений. Однако эти бабочки не могут пережить холодную зиму. Каждую осень громадные стаи бабочек, подобно птицам, летят на юг до Мексики или еще дальше, где в количестве от нескольких тысяч до миллионов особей скапливаются на отдельных деревьях. В это время имеет место задержка развития половых продуктов (репродуктивная диапауза). Однако уже в феврале бабочки спариваются и начинают обратный перелет на север (M.J.Tauber et al., 1986). Осенняя миграция в южном направлении, но не столь дальняя, наблюдалась и у ряда других бабочек и даже клопов.
Сезонный покой
Во время неблагоприятного сезона насекомые прекращают свою активность и, как правило, не размножаются. Это происходит не только потому, что при низкой температуре насекомые не могут двигаться, так как состояние покоя может быть и летним. Насекомому необходимо дожить до конца неблагоприятного сезона, что невозможно без экономии энергетических ресурсов и воды. Соответственно жизненные циклы насекомых помимо активной жизнедеятельности (роста, развития, питания, размножения, миграций) могут включать в себя при необходимости и временное состояние сезонного покоя.
Сезонный покой может быть двух типов: экзогенный и эндогенный (криптобиоз) (Р.С.Ушатинская, 1990). В первом случае насекомое прекращает свою активность под непосредственным воздействием высоких или низких температур, а также, например при пересыхании водоема, в котором оно обитает. Во втором – насекомое на основе восприятия сигнальных изменении внешней среды заранее приспосабливается к неблагоприятным условиям, подгоняя к этому времени определенную стадию развития и осуществляя определенную физиологическую перестройку организма.
Экзогенный покой типичен для насекомых южного происхождения, таких, как, например, луговая (Mythimna unipuncta Hw.) и малая наземная (Laphygma exigua Hb.) совки. Обитание этих бабочек в умеренной зоне, по-видимому, связано с их высокой миграционной способностью. Экзогенный покой имеет место также у ряда тлей, способных перезимовывать на любой стадии развития. Он возможен и у некоторых арктических насекомых, где низкие температуры могут быть в любое время года и устойчивость к ним должна быть "хронической". В случае экзогенного покоя насекомые приходят в активное состояние сразу же после окончания неблагоприятного воздействия, что особенно важно в арктической обстановке.
Гораздо чаще у насекомых наблюдается предварительная подготовка к неблагоприятным условиям и соответственно эндогенный покой. В этом случае насекомому необходимо прежде всего согласовать свое развитие с сезонностью таким образом, чтобы на неблагоприятный сезон пришлась стадия с наибольшей устойчивостью. Это достигается при помощи так называемых количественных реакций, прежде всего ускорения или замедления развития отдельных стадий. Насекомое должно также уловить изменения внешней среды, предшествующие неблагоприятному сезону, и, восприняв эти сигналы, изменить свое поведение так, чтобы заранее найти место, где легче всего перенести неблагоприятные условия, накопить запас резервных веществ в теле и, наконец, перестроить свои физиологические процессы таким образом, чтобы организм был максимально устойчив к неблагоприятным воздействиям.
Диапауза
Как уже отмечалось выше, ряд насекомых, особенно примитивных, способен переносить замораживание или высушивание и сохранять при этом жизнеспособность. Большинство же насекомых приурочивает к неблагоприятному сезону особое физиологическое состояние – диапаузу. Насекомые в состоянии диапаузы устойчивы не только к холоду, но и ко многим другим неблагоприятным воздействиям (С.И.Черныш, 1990). Диапауза – не обязательно является приспособлением к сезонности, но может возникать, например, при повышенной плотности популяции.
Наиболее характерной для любой диапаузы является приостановка или существенное замедление развития либо самого насекомого, либо его гонад, если диапаузирует имаго. Диапаузирующие насекомые не питаются или питаются очень редко, их спаривание никогда не наблюдается. Отметим, что остановка морфогенеза и снижение уровня обменных процессов не означают, что во время диапаузы не происходит вообще никаких физиологических процессов. Иногда говорят даже о диапаузном развитии или о процессе изживания диапаузы.
Очевидно, что у тропических и некоторых субтропических видов диапауза может вообще отсутствовать. Если растение пригодно для питания фитофагов не круглогодично, насекомые на неблагоприятное время переходят на другие, иногда менее подходящие для них растения. Даже на севере Англии развитие и размножение некоторых видов дрозофил может проходить круглогодично, хотя зимой развитие замедлено и увеличивается объем жирового тела.
Дата добавления: 2016-06-29; просмотров: 1801;
Поиск по сайту
Узнать еще
- I.1.1 ПРЕДМЕТ МОЛЕКУЛЯРНОЙ ФИЗИКИ
- Активные – воздействуют на предмет труда, перемещают его, контролируют ход производства (машины, оборудование, т/с, инструмент)
- Актуальность изучения экологии
- Алфавитно-предметный указатель
- Аналитическая химия и химический анализ. Предмет и задачи аналитической химии. Классификация методов химического анализа.
- Аналогично работают ПТК СВШД и системы обнаружения случайных предметов (СОСП).
- Анатомо-физиологические аспекты потребностей человека. Задачи изучаемого предмета.
- Антропогенное влияние на природу. Проблема экологии.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
