Предмет экологии насекомых 4 глава
Следовательно, насекомые имеют большие возможности, перемещаясь в пределах биотопа или зарываясь в почву, находить для себя наиболее благоприятные условия. Соответственно, и температура, с которой реально имеет дело насекомое, может существенно отличаться от обычных метеорологических показателей. Температура оказывает существенное влияние на поведение насекомых, накладывает определенные рамки на их способность выжить, видоизменяет морфологию и окраску особей.
Измерения температуры и термостатирование.
Обычные измерения температуры производятся с помощью термометров разных типов. Измерения температуры недопустимо производить при прямом солнечном освещении самого термометра, так как в этом случае он показывает не столько температуру воздуха, сколько степень его нагрева солнечными лучами. В полевых условиях удобно измерять температуру с помощью психрометра Ассмана, который служит и для измерения влажности воздуха, а также имеет отражающую солнечные лучи металлическую изоляцию. Однако и этот прибор может при измерении находиться на прямом солнечном свете не более 2–3 мин.
Микроклиматические измерения удобно проводить с помощью электронного термометра с щупом, позволяющим проникать в укрытия. При отсутствии такого термометра используют термосопротивления, которые можно с помощью жесткого резинового шланга вводить, например, глубоко в норы. При измерении температуры тела насекомого часто использовались, так называемые, термопары. Термопара – это две проволоки из разных сплавов (например, константана и меди), концы которых спаяны вместе таким образом, что на схеме образуют замкнутый прямоугольник или круг. Если эти спаи держать при разной температуре, то возникает ЭДС, которую можно измерить с помощью гальванометра. На практике обычно один спай погружают в термос с мелко наколотым тающим льдом (0° С), а другой вводят в то микропространство, температуру которого надо измерить. Предварительно экспериментальным путем составляют шкалу, показывающую соотношение между показаниями гальванометра и разностью температур, при которых находятся спаи.
Для создания строго контролируемой температуры используют термостаты. При этом следует помнить, что обычный термостат, имеющий только нагреватель, не может создать температуру ниже температуры воздуха в самой лаборатории. Поэтому в летнее время часто температура внутри термостата оказывается выше той температуры, на которую он был настроен. Дня энтомологических целей гораздо удобнее пользоваться особыми термостатами, имеющими помимо нагревателя также и холодильное устройство. Такие термостаты независимо от температуры воздуха в лаборатории могут создать любую температуру в пределах от +6 до +40° С. Особенно велика потребность в такой технике, если условия опыта требуют специального освещения камеры термостата, так как практически любой источник света создает дополнительный нагрев.
Любые термостаты не гарантируют полного равенства температур во всех точках их внутреннего пространства, поэтому контрольные термометры должны находиться своими резервуарами возможно ближе к камерам с насекомыми. Равномерность нагрева внутри термостата увеличивается, если там находится небольшой вентилятор, однако при этом надо быть уверенным, что звук и вибрация, создаваемые им, не влияют на подопытных насекомых.
Выпускались также климокамеры, создающие возможность контроля сразу за многими факторами среды, однако технически они гораздо сложнее в обслуживании и занимают много места. Иногда термостатируют целиком все лабораторное помещение.
Влияние температуры на поведение насекомых
Температура может оказывать двоякое воздействие на насекомых. Во-первых, с ее повышением и соответственным увеличением скорости обмена веществ появляется возможность того или иного рода поведения, а также увеличивается скорость движения (непосредственное воздействие). Во–вторых, температура среды и ее изменения воспринимаются терморецепторами, при этом насекомое может активно искать благоприятную для себя зону и уходить из неблагоприятной (сигнальное воздействие). Как слишком низкие, так и слишком высокие температуры подавляют активность.
Соответственно, температура может почти полностью определять поведение насекомых в арктических условиях, где активность возможна лишь в течение короткого интервала времени, в умеренной зоне – зимой, весной и поздней осенью, в отдельные дни и летом, а в пустынной зоне – почти ежедневно в летнее время.
Диапазон температур, в пределах которого возможна активность, конечно, различен для разных видов. Клопы, жуки, цикадки и крылатые муравьи более чувствительны к понижению температуры, чем двукрылые и бабочки. В целом насекомые, летающие летом, гораздо менее устойчивы к низким температурам, чем весенние и осенние. Лет ранневесенних видов не останавливает даже 0° С. Высокая температура в середине дня заметно снижает активность многих насекомых. В целом, кривая зависимости скорости движения от температуры, полученная на примере поведения рабочих муравьев, отдаленно напоминает кривую нормального распределения – как высокие, так и низкие температуры подавляют активность. Несколько иначе выглядит кривая зависимости процента подвижных особей от температуры. Здесь этот процент резко возрастает при достижении температурного порога, а далее остается неизменным для довольно большого диапазона температур.
Разные типы подвижности и активности насекомых ограничены различными температурными диапазонами. Так, полет допустим в пределах более узкого температурного интервала, чем ползание. Тип полета тоже зависит от температуры воздуха. Движение крыльев бобовой тли может наблюдаться с 6,5° С, горизонтальный полет при сбрасывании с высоты – с 13° С, а активный полет – с 15° С (A.J.Cockbain, 1961).
Особенно четко зависит от температуры возможность спаривания насекомых. Интересно, что у многих мух спаривание имеет место только после энергичных и продолжительных движений, особенно в прохладную погоду. Поэтому не исключено, что "брачные танцы" насекомых могут иметь, помимо этологического, и чисто физиологическое значение.
Насекомые активно ищут участки с наиболее благоприятной для них температурой. Яркий пример связанных с температурой ежесуточных вертикальных миграций дает поведение большой хлопковой тли Acyrthosiphon gossypii Mordv. на растениях хлопчатника (В.Б.Чернышев и др., 1981). Поздним утром и в начале дня тли сидят на освещенных солнцем верхушечных листьях растения. В середине дня они спускаются по растению вниз, где значительно прохладнее. Часть тлей в это время прячется в трещинах почвы около растения. Позже к вечеру они переходят на стебель и постепенно перемещаются вверх. Некоторые тли опять переходят на верхушечные освещенные солнцем листья. Ночью они располагаются, в основном, на стебле, а ранним утром, когда холоднее всего, до 80% тлей концентрируется ввиде грозди около точки роста – наиболее теплой части растения. В результате таких перемещений суточные изменения температуры для тлей существенно сглаживаются. Днем температура, при которой они находятся, ниже температуры воздуха на 10–15° , а ночью – выше на 2–3° . Подобные перемещения по растениям описаны и для многих других насекомых.
Температура тела насекомых может существенно отличаться от температуры воздуха в результате физиологических процессов. Благодаря мышечным движениям тело насекомого может нагреваться. Так, по данным И. Д Стрельникова (1940), температура тела летающих бабочек, мух и перепончатокрылых всегда около 35–40° и мало зависит от температуры окружающего воздуха. Сохранению тепла у ночных насекомых может способствовать густой волосяной покров. Но даже и у неподвижных живых насекомых из–за обменных процессов температура тела выше окружающей на 0,1–0,5°. Другая причина повышения температуры тела насекомых – это их нагрев в солнечных лучах, особенно резко проявляющийся в высокогорной местности. На низменности отмечен дополнительный нагрев тела насекомого немногим более 5–6°, в горах эта разность температур может достигать 30°. Насекомые способны активно изменять уровень нагрева тела, принимая разные позы по отношению к солнечным лучам или раскрывая крылья, служащие отличным приемником тепла дневные бабочки принимают широко известную красивую позу с распростертыми крыльями только в относительно прохладную погоду, в жаркую же погоду крылья захлопываются над спинкой сразу же после посадки.
Очень большое значение в регуляции температуры тела насекомых имеет их окраска. Типичные для многих дневных насекомых металлические окраски отражают излишнюю солнечную радиацию. Ночью особенно неблагоприятны темные окраски, так как они способствуют потере тепла путем радиации. Поэтому в среднем ночные насекомые заметно светлее, - чем дневные.
Влияние на насекомых низких и высоких температур
Жизнь любого насекомого возможна только в определенном температурном интервале. Р.С.Ушатинская (1957) в этом отношении различает шесть следующих температурных зон:
1. Зона активной жизни лежит в среднем в пределах от 3 до 40°. Примерно в середине этой зоны – температуры, обеспечивающие минимальную смертность и максимальную плодовитость насекомых.
2 Нижняя зона переживания (зона холодового оцепенения). Здесь возможность выжить зависит от уровня температуры, продолжительности ее воздействия и, конечно, от вида насекомого.
3. Нижняя смертельная (летальная) зона, в которой происходит замерзание и кристаллизация жидкостей тела, а также повреждение кристаллами протоплазмы клеток. Эти необратимые изменения несколько различны у разных видов.
4. Зона витрификации, в которой жидкость, вместо того, чтобы образовывать кристаллы, становится витрифицированной, т.е. подобной стеклу. Витрификация возможна далеко не у всех насекомых. При этом происходит приостановка всех жизненных процессов, аналогичная анабиозу. В таком состоянии насекомое может вынести охлаждение почти до абсолютного нуля. Однако витрифицированная жидкость неустойчива и может постепенно кристаллизоваться.
5. Верхняя зона переживания (зона теплового шока). Так же как и в нижней зоне переживания продолжительность жизни насекомых зависит от уровня температуры и длительности ее воздействия.
6. Верхняя смертельная зона, в которой происходят необратимые явления: коагуляция белков и инактивация ферментов.
Влияние температуры на насекомое во многом зависит от его вида и образа жизни. Насекомые, обитающие в умеренной, а тем более в полярной зоне, наиболее устойчивы к низким температурам. Многие из этих насекомых легко переносят многократное замерзание и оттаивание. В умеренной зоне устойчивость насекомых к холоду закономерно изменяется в зависимости от сезона и наиболее высока в середине зимы, при этом наиболее устойчивыми оказываются насекомые, зимующие не под снежным покровом, а под корой деревьев и в пустых стеблях растений. Холодостойкость водных насекомых относительно мала в любое время года.
Если охлаждение не является очень глубоким и наступило внезапно, насекомое впадает в состояние холодового оцепенения. При повышении температуры такое насекомое быстро становится активным. В оцепеневшем состоянии насекомые могут без особого вреда для себя находиться от нескольких дней до недель. Временное охлаждение, задерживающее развитие и существенно удлиняющее жизнь насекомого, часто используется энтомологами в практической работе. Обычная температура холодильника (+2 – +4°) вполне достаточна для хранения насекомых. Следует помнить, что насекомые при таком хранении погибают не столько от холода, сколько от высушивания. Поэтому рекомендуется садок с насекомыми помещать в полиэтиленовый плотно закрытый пакет с куском ваты, смоченной водой.
В природе холода наступают в определенное время года, и перед наступлением неблагоприятного сезона в организме насекомого происходят иногда очень глубокие физиологические перестройки. Они связаны с определенным физиологическим состоянием – диапаузой.
Что происходит с насекомыми при дальнейшем понижении температуры? Появление ледяных кристаллов в клетках тканей насекомого, по–видимому, для него всегда смертельно. Тем не менее ряд насекомых способен переносить морозы в течение длительного времени. Таких насекомых можно разделить на две категории (M.J.Tauber et al., 1986): устойчивые к замерзанию (после замерзания внеклеточной жидкости они остаются живыми) и неустойчивые (гибнущие после замерзания, но имеющие специальные приспособления, чтобы ему противостоять). По–видимому, в редких случаях возможно и сочетание устойчивости к замерзанию с механизмами, препятствующими замерзанию. Явление витрификации, если и встречается, бывает сравнительно редко, и само это состояние воды достаточно неустойчиво.
Устойчивость к замерзанию не встречается среди филогенетически низших групп насекомых, а также среди многоножек, скорпионов и пауков. Такую устойчивость выработали лишь некоторые двукрылые, бабочки, жуки и сетчатокрылые, и то лишь на определенной стадии развития. У этих насекомых жидкости тела замерзают при относительно высокой для насекомых температуре – не ниже –10°С. Особенностью этих насекомых является наличие в гемолимфе особых белковоподобных веществ, способствующих образованию кристаллов льда между органами. Эти кристаллы притягивают к себе молекулы воды, оставшиеся свободными. По–видимому, при этом резко снижается возможность кристаллизации воды внутри клеток. К числу таких устойчивых к замерзанию насекомых можно отнести, например, бабочку–махаона, куколка которого в замороженном состоянии способна переносить температуру до –196° С.
Гораздо более распространена среди насекомых способность противостоять замерзанию. Такие насекомые вырабатывают специальные приспособления, суть действия которых сводится к снижению точки переохлаждения, а также к максимальному удалению веществ, способствующих образованию кристаллов льда по крайней мере внутри клеток. Для таких насекомых при их обитании в умеренной зоне точка переохлаждения, ниже которой возможно замерзание, лежит ниже –30° С, а для насекомых арктической зоны – ниже –60° С.
Каковы реальные возможности повышения холодостойкости у насекомых?
Во-первых, это уменьшение общего количества воды в теле и связывание ее коллоидами. О значении воды для холодостойкости свидетельствует такой факт. Зимующие гусеницы златогузки содержат в теле до 69% воды и выдерживают температуру –14° до 158 дней. Активные же гусеницы в летнее время содержат более 80% воды и могут переносить ту же низкую температуру не более 1,5 –4 ч.
Во-вторых, это увеличение содержания жира. Так, у тех же зимующих гусениц златогузки содержание жира доходит до 6%, в то время как летом оно примерно 4%.
В-третьих, это увеличение количества резервных углеводов, особенно гликогена, являющегося гидрофильным коллоидом.
В-четвертых, это повышение концентрации различных веществ, растворенных в жидкостях тела. Известно, что 1 моль любого вещества на 1 литр раствора понижает температуру замерзания последнего почти на 2° Эффект от нескольких веществ, находящихся в жидкости, суммируется.
Среди таких веществ можно назвать некоторые сахара (трегалоза, глюкоза, фруктоза), специальные белки и аминокислоты. Кроме того, в теле зимующих насекомых нередко в большом количестве (до 25% от массы тела) присутствует широко используемый в технике антифриз – глицерин или аналогичные ему по действию вещества. Глицерин здесь обычно не является только пассивным антифризом и определенным образом распределяется в теле насекомого. Во всяком случае, искусственная инъекция глицерина не всегда приводит к повышению холодоустойчивости. После окончания зимовки глицерин превращается в гликоген.
Необходимость удаления из тела веществ, способствующих появлению кристаллов, приводит иногда к определенным изменениям пищевой диеты. У насекомого, полностью готового к зимовке, кишечник освобождается от содержимого.
Рекорд холодостойкости поставили личинки одного из видов комаров–дергунов, обитающие в горных районах Африки. Эти личинки развиваются во временных водоемах – углублениях скал, заполняемых водой во время дождей. Все эти водоемы вместе с личинками быстро высыхают, но личинки в сухом виде остаются живыми и после увлажнения опять приходят в активное состояние. Личинки в сухом виде без какого–либо ущерба выдерживают температуру почти абсолютного нуля (–270°).
Гораздо сложнее для насекомых противостоять высоким температурам, которые быстро приводят к нарушениям метаболизма, коагуляции белков и гибели. Какое–то время насекомые способны поддерживать температуру тела ниже температуры окружающего воздуха за счет испарения влаги, которое усиливается в результате разрушения высокой температурой воскоподобной оболочки тела. Естественно, что этот эффект определяется влажностью воздуха. Так, черные тараканы во влажном воздухе быстро погибают при +38°, а в сухом, обеспечивающем испарение, выживают какое–то время и при +48°. В некоторых случаях температура тела может быть понижена размазыванием по телу капель жидкости, выделяемых из ротового или анального отверстий (С.В.Томс, 1981).
Результатом специальных физиологических приспособлений является способность некоторых насекомых жить в горячих вулканических источниках при температуре воды до +65°. Таковы личинки некоторых мух–прибрежниц и львинок. Полной им противоположностью являются насекомые, активные на снегу в зимнее время. Для них губительна комнатная температура (+20°). Сходным образом температура +35° за минуты убивает обитающих в пещере при постоянной температуре +11° кузнечиков.
Влияние температуры на развитие насекомых.
Продолжительность развития (N) от момента откладки яиц и до появления имаго или каждой отдельной стадии обычно измеряется днями. На практике часто применяется понятие скорости развития (V). Очевидно, что V=1/N.
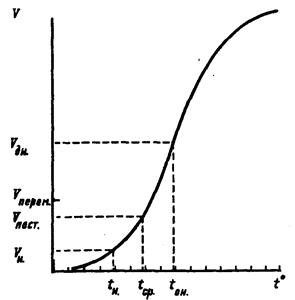
Экспериментальные данные показывают (рис. 2), что с увеличением температуры продолжительность развития постепенно уменьшается, при определенном диапазоне температур достигает минимума, а с дальнейшим повышением температуры может опять увеличиваться. В целом такая кривая очень напоминает цепочку, подвешенную за два конца. Правая часть кривой, показывающая замедление развития при высоких температурах, часто не реализуется из-за высокой смертности насекомых.
Скорость развития, соответственно, сначала медленно возрастает с повышением температуры. Эта скорость увеличивается все быстрее вплоть до точки перегиба кривой, затем кривая становится более пологой, доходит до максимума, а потом либо остается на прежнем уровне, либо несколько снижается при особо высоких температурах.
Очевидно, что зависимость скорости развития от температуры должна в значительной степени объясняться обычными физико-химическими закономерностями. Согласно правилу Вант-Гоффа, скорость химического процесса с повышением температуры на 10° увеличивается в 2 – 3 раза. Тогда, определив скорость развития Vo при какой-либо температуре 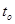 , можно рассчитать скорость развития при другой температуре t по следующей формуле:
, можно рассчитать скорость развития при другой температуре t по следующей формуле:
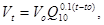
где 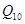 – коэффициент увеличения скорости.
– коэффициент увеличения скорости.
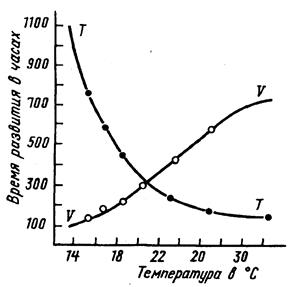

Рис.2. Продолжительность развития куколок большого мучного хрущака Tenebrio molitor (Т) и скорость их развития (V) в зависимости от температуры (по А.Крогу из В.В.Яхонтова, 1969)
Таким образом, уравнение Вант-Гоффа – это показательная (экспоненциальная) функция. Очевидно, что оно пригодно только для описания начальной, резко восходящей части экспериментальной кривой. Оно плохо описывает также и ход процесса в области низких температур. Наиболее точно отражает весь ход кривой зависимости скорости развития насекомого от температуры следующее эмпирическое уравнение:
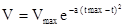
где 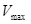 – максимально возможная скорость развития;
– максимально возможная скорость развития; 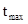 - температура, при которой достигается эта скорость; а – коэффициент скорости изменения V относительно t.
- температура, при которой достигается эта скорость; а – коэффициент скорости изменения V относительно t.
В энтомологической практике иногда используют также формулу логистической функции, которая не описывает спад скорости развития при высоких температурах:
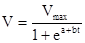 ,
,
где а и b – константы уравнения, определяющие наклон и точку перегиба кривой.
Очень удобна для расчетов, хотя и неточна, так называемая сумма эффективных температур. Очевидно, что всегда имеет место пороговая температура развития, ниже которой насекомое не может развиваться (t 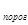 ). Если выращивать насекомое при какой-то постоянной температуре t, которая выше пороговой, то эффективная температура (
). Если выращивать насекомое при какой-то постоянной температуре t, которая выше пороговой, то эффективная температура ( 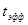 ) равна:
) равна:
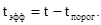
Правило суммы эффективных температур предполагает, что для развития каждого насекомого требуется определенная доза тепла, т.е. определенное число "градусо-дней", или тепловая постоянная (К). Тогда при выращивании насекомых в условиях постоянной температуры t она равна:
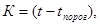
где N – срок развития в днях. Если температура, как в природе, день ото дня меняется, то формула тепловой постоянной принимает следующий вид:
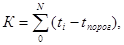
где t – средняя суточная температура для каждого из дней, i – знак суммы.
Чтобы найти пороговую температуру, не обязательно проводить сложные эксперименты по развитию насекомых при низких температурах. Достаточно определение сроков развития для двух уровней температуры 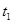 и
и 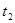 . Тогда пороговую температуру можно высчитать по формуле:
. Тогда пороговую температуру можно высчитать по формуле:
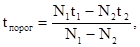
где N и N соответствующие сроки развития. Обычно пороговая температура для развития насекомых лежит в пределах от +3° до +8° С.
Зная тепловую постоянную К, можно рассчитать срок развития при любой температуре:
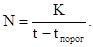
Эта формула соответствует гиперболической зависимости типа 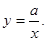
Следовательно, кривая зависимости срока развития от температуры будет иметь вид гиперболы. Однако эта гипербола не может отразить увеличение срока развития при высоких температурах.
Скорость развития согласно этой формуле будет иметь вид
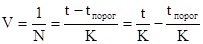
так как 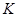 и
и 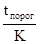 величины постоянные, то эта формула соответствует прямолинейной зависимости типа
величины постоянные, то эта формула соответствует прямолинейной зависимости типа
у = ах+ b .
Таким образом, формула суммы эффективных температур применима только в зоне, близкой к точке перегиба восходящей части кривой зависимости скорости развития от температуры. При более низких и более высоких значениях температуры, где кривая уплощается, эта формула дает менее точные результаты.
Сравнивая тепловую постоянную с суммой эффективных температур за сезон, можно приближенно определить возможное количество поколений данного вида за это время. Здесь сумма эффективных температур за сезон (Э) высчитывается почти по той же формуле, что и тепловая постоянная, но вместо срока развития N, здесь стоит число дней в сезоне (n), когда температура превышает пороговую:
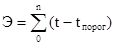
Если, например, сумма эффективных температур в течение вегетационного сезона равна 3200°, а тепловая постоянная – 1000°, то это означает, что в данной местности за год могут развиться три поколения данного вида.
Как уже отмечалось выше, формула суммы эффективных температур довольно приближенно отражает реальную зависимость. Имеются и другие причины, затрудняющие использование этой формулы.
Во-первых, температура тех микроучастков, где находятся насекомые, может отличаться от метеорологических измерений на 10 и более градусов (например, на поверхности растения).
Во-вторых, температура тела насекомого может существенно отличаться от температуры воздуха из-за метаболического тепла и дополнительного нагрева тела в солнечных лучах.
В-третьих, есть основания предполагать, что средняя сумма эффективных температур различна для популяции при нарастании ее численности (начало вспышки массового размножения) и при спаде (кризис). В первом случае развитие идет быстрее и сумма эффективных температур может быть меньше.
В-четвертых, в природе температура все время меняется. Как показывают эксперименты, влияние таких переменных температур не сводится к простой суммации, а представляет собой довольно сложное адаптационное явление. Вопрос о влиянии температурных циклов на насекомых интересен не только с экологической точки зрения, он очень важен для практики массового разведения насекомых. Поэтому остановимся на нем несколько подробнее.
Согласно большинству публикаций, срок развития насекомых заметно укорачивается при переменной температуре по сравнению с соответствующей средней постоянной температурой. Однако эффект явно зависит от вида насекомого. Если, например, у трихограммы при переменной температуре развитие заметно ускоряется, то у дрозофил, наоборот, происходит замедление развития. При переменной температуре имеет место не только ускорение развития, но может резко уменьшаться и смертность, особенно на стадии яйца. Развитие некоторых жужелиц, возможно только при переменной температуре. У ряда бабочек и жуков при переменной температуре заметно возрастает плодовитость.
Все эти эффекты зависят также и от параметров терморитма: от длины "температурного дня", от резкости изменений температуры, от уровней максимальной и минимальной температуры. По-видимому, желательно, чтобы изменения температуры были постепенными, при резких сменах температур может существенно снизиться жизнеспособность насекомых из-за температурного шока или их залипания в результате запотевания стенок садка. Слишком высокий верхний уровень температуры может привести к гибели или стерильности насекомых. Нижний уровень температуры в цикле не столь ограничен и может быть выше или ниже порога развития.
Возможны несколько механизмов, лежащих в основе действия переменных температур. Во-первых, особенности влияния переменных температур можно объяснять тем (Н.И.Горышин), что кривая (рис. 3), описывающая зависимость скорости развития от температуры, имеет изгибы как в нижней, так и верхней части. Если средний уровень переменной температуры ниже точки перегиба кривой, то такая переменная температура должна ускорять развитие, потому что снижение температуры ниже средней для всего цикла мало что изменит в скорости развития, а ее повышение приведет к резкому возрастанию скорости развития. Наоборот, при высоких средних переменная температура должна замедлять развитие, так как нижняя – будет попадать в область резкого изменения функции, а высокая – либо почти не увеличит скорость развития, либо еще ее уменьшит.
 Во-вторых, ускорение развития под влиянием переменной температуры, равно как и увеличение жизнеспособности, можно объяснить особенностями физиологической адаптации организма к изменениям температуры. Повышение температуры приводит к резкому "всплеску" метаболизма, который затем постепенно снижается до уровня, типичного для данной высокой температуры.
Во-вторых, ускорение развития под влиянием переменной температуры, равно как и увеличение жизнеспособности, можно объяснить особенностями физиологической адаптации организма к изменениям температуры. Повышение температуры приводит к резкому "всплеску" метаболизма, который затем постепенно снижается до уровня, типичного для данной высокой температуры.
Рис.3. Скорость развития насекомого при постоянной и переменной температуре (продолжительности термо– и криофазы одинаковы и равны 12 ч) V– скорость развития, t – температура, t  – температура в течение криофазы,
– температура в течение криофазы, 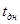 – температура в течение термофазы,
– температура в течение термофазы, 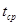 – средняя температура цикла,
– средняя температура цикла, 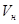 – скорость развития во время криофазы,
– скорость развития во время криофазы, 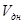 – скорость развития во время термофазы,
– скорость развития во время термофазы, 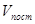 – скорость развития при постоянной температуре, равной средней температуре цикла,
– скорость развития при постоянной температуре, равной средней температуре цикла, 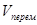 – реальная скорость развития при переменной температуре
– реальная скорость развития при переменной температуре
Понижение температуры, наоборот, приводит к временному резкому снижению обмена, которое, однако, меньше по амплитуде, чем всплеск, и не может его полностью компенсировать. Тем не менее вопрос о механизме влияния переменной температуры на насекомых не может считаться окончательно решенным.
Дата добавления: 2016-06-29; просмотров: 1684;
Поиск по сайту
Узнать еще
- I.1.1 ПРЕДМЕТ МОЛЕКУЛЯРНОЙ ФИЗИКИ
- Активные – воздействуют на предмет труда, перемещают его, контролируют ход производства (машины, оборудование, т/с, инструмент)
- Актуальность изучения экологии
- Алфавитно-предметный указатель
- Аналитическая химия и химический анализ. Предмет и задачи аналитической химии. Классификация методов химического анализа.
- Аналогично работают ПТК СВШД и системы обнаружения случайных предметов (СОСП).
- Анатомо-физиологические аспекты потребностей человека. Задачи изучаемого предмета.
- Антропогенное влияние на природу. Проблема экологии.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
