Популяционные, экологические и эволюционные проблемы трофологии. Биосфера как трофосфера
С деятельностью живых систем связана та часть поверхности Земли, которая объединена под названием биосферы. Биосфера, являющаяся самой крупной экосистемой, представляет собой открытую систему, т. е. получает и отдает энергию. Поток энергии направлен в одну сторону, причем часть поступающей солнечной энергии трансформируется в органическое вещество, а большая часть деградирует. Энергия может накапливаться, а затем снова освобождаться и экспортироваться. В отличие от энергии пищевые вещества, в том числе витальные биогенные элементы (углерод, азот, фосфор и т. д.) и вода, используются многократно (обзор: Odum, 1986).
Ежегодно фотосинтезирующие организмы продуцируют до 1017 т (около 100 млрд. т) органического вещества. За это же время весьма значительное количество вещества разрушается и превращается в углекислый газ и воду в результате дыхания растений (Vallentyne, 1962). Предполагается, что преобладание органического синтеза над дыханием явилось главной причиной уменьшения содержания углекислого газа в атмосфере и накопления в ней кислорода, уровень которого в настоящее время достаточно высок. Именно это и способствовало эволюции и развитию высших форм жизни на Земле (обзор:Odum, 1986).
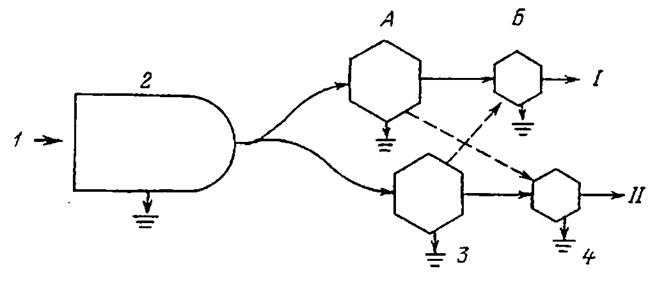
| Рис. 1.6. Модель потока энергии, показывающая связь между пастбищной и детритной трофическими копями (по:Odum, 1986). I - пастбищная трофическая цепь; II - детритная трофическая цепь. А - растительноядные, Б - хищные животные. 1 - солнечный свет; 2 - растения; 3 - потребители детрита; 4 - хищники. |
Перенос энергии пищи от ее источника - аутотрофов (растений) через ряд организмов путем поедания одних организмов другими составляет трофическую цепь. При каждом очередном переносе до 80-90% потенциальной энергии переходит в тепло. В связи с этим чем короче трофическая цепь, тем большее количество энергии доступно для популяции.
Формально трофические цепи можно разделить на пастбищную, начинающуюся от растений и идущую к растительноядным животным, и детритную, которая идет от мертвого органического вещества к микроорганизмам, а затем к детрофагам и хищникам (рис. 1.6). Важно, что трофические цепи не изолированы, а тесно переплетены, образуя так называемые трофические сети.
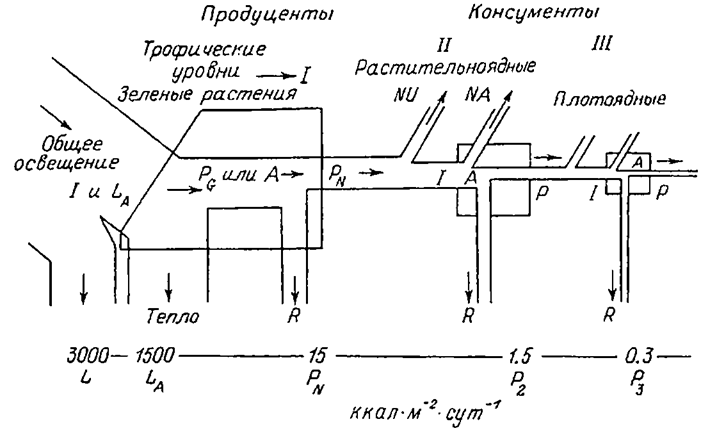
| Рис. 1.7. Упрощенная схема потока энергии на трех трофических уровнях в линейной трофической цепи (по: Odum, 1986). I, II, III - трофические уровни. I -общее поступление энергии; LA - свет, поглощаемый растительным покровом; PG - валовая первичная продукция; А -общая ассимиляция; PN - чистая первичная продукция; Р - вторичная продукция консументов; NU-неиспользуемая (накапливаемая или экспортируемая) энергия; NA-неассимилированная консументами (выделенная с экскрементами) энергия; R- дыхание. Цифры внизу - порядок величины потерь энергии при каждом переносе, начиная с поступления солнечного излучения в количестве 3000 ккал.м-2сут-1. |
Считается, что организмы, получающие энергию от Солнца через одно и то же количество этапов, принадлежат к одному уровню. При этом популяция может занимать не только один, а несколько уровней в зависимости от используемых источников энергии. Условно можно выделить 4 трофических уровня: 1) продуценты (зеленые растения); 2) первичные консументы (травоядные);3) вторичные консументы (первичные хищники, поедающие травоядных);4) третичныеконсументы (вторичные хищники). Принято, что человек чаще всего одновременно является первичным и вторичным консументом, так как в его диету, как правило, входит пища как растительного, так и животного происхождения. В этом случае поток энергии разделяется между этими трофическими уровнями в соответствии с долями растительной и животной пищи. В превосходном обзоре Ю. Одума (Odum, 1986) представлена упрощенная схема потока энергии на трех уровнях (рис. 1.7). Как можно видеть, поток энергии через трофический уровень соответствует общей ассимиляции (А) на этом уровне, а последняяравна продукции (Р) биомассы плюс дыхание (R). На каждом следующем трофическом уровне поток энергии уменьшается. На первом трофическом уровне поглощается до 50% падающего света, а в энергию пищи трансформируется только 1% поглощенной энергии. На двух следующих уровнях вторичная продуктивность составляет лишь 10% от предыдущей. В целом средняя эффективность переноса энергии между трофическими уровнями составляет 20% и менее. Существует предположение, что регуляция первичной продукции зависит от числа звеньев в трофической цепи (Smith, 1969).
Что касается трофических цепей, то в большинстве случаев в каждом следующем звене поток доступной энергии уменьшается на один порядок. Равновесие между синтезом и деструкцией веществ - необходимое условие поддержания жизни в планетарном масштабе и существования каждого вида в отдельности.
Живые организмы, представляющие собой активную силу, действующую сегодня, составляют лишь сравнительно небольшую часть биосферы. Поэтому хотя носителями жизни являются отдельные организмы, жизнь, согласно В. И. Вернадскому (1926,1965,1980), возможна лишь как планетарное явление, как форма существования биосферы с обязательным для нее круговоротом веществ и потоков энергии, так называемым биотическим круговоротом. Этот круговорот определяется как метаболическими процессами, происходящими в отдельных организмах, так и связями между ними. Связи между различными организмами также в значительной мере являются трофическими.
Биотический круговорот в целом можно рассматривать как трофический процесс, а сами организмы включены в трофические цепи, точнее трофические сети, где каждый вид использует определенные источники питания и вместе с тем сам служит пищевым объектом. Источники питания, количество, свойства и доступность пищи в значительной мере лимитирует распределение и численность любой популяции, а также во многом определяют ее эволюционную судьбу. Следовательно, одним из условий устойчивости биосферы является биотический круговорот, или цикл, в который как важный компонент входят трофические взаимодействия. Последние обеспечивают образование биологической массы, ее миграцию и модификацию, а также постепенную деградацию до неорганических элементов, которые вновь включаются в начальные звенья цикла. Многие неорганические вещества превращаются в органические и трансформируются в метаболических звеньях трофических цепей, а затем в идеальных случаях возвращаются в метаболические звенья компонентов биотического круговорота. (Закономерности круговорота веществ и энергии в биосфере, проблема трофических связей и их роли в биосфере, ее эволюция и ряд других вопросов детально рассмотрены в ряде обобщающих сводок: Вернадский, 1926, 1965, 1980; Биосфера, 1972; Будыко, 1984; Уголев, 1985, 1986а, 1987а; Аксенов, 1986; Добровольский, 1986; Соколов, 1986; Allen, Nelson, 1986; Odum, 1986; Лапо, 1987; Greenetal., 1990, и др.)
Таким образом, трофические связи являются обязательным элементом жизни как биосферного явления, а проблема происхождения жизни на Земле в конечном итоге сводится к вопросу об образовании первичной биосферы с ее первичными трофическими взаимоотношениями (см. гл. 9). Этот вопрос был поставлен еще В. И. Вернадским, как и вопрос о трофических связях в современной биосфере.
Понимание биосферы как трофосферы (Уголев, 1986а), состоящей из различных трофоценозов с их цепными и разветвленными связями, обеспечивающих циркуляцию веществ и энергии, позволяет решать проблемы охраны окружающей среды и поддерживать экологическое равновесие на основе анализа трофических соотношений и их сохранения. В некоторых случаях при нарушении трофических цепей возможно их восстановление за счет включения недостающих звеньев.
Следует обратить внимание на несколько аксиоматических положений: 1) в основе энергетики жизни и образования органических веществ лежат преимущественно процессы фотосинтеза, которые осуществляются за счет солнечной энергии; 2) основная часть энергии расходуется в результате метаболизма самими аутотрофными организмами; 3) лишь небольшая часть аккумулированного материала (примерно 10%) переходит в следующее звено трофической цепи,и т. д. Следовательно, как уже отмечено выше, лишь небольшая часть энергии, накопленной в органических веществах, передается по трофическим цепям. Однако именно эта неметаболизированная часть и является самой важной, так как она определяет биологический состав биосферы, ее единство, гомеостаз и многие другие важнейшие свойства (обзор: Odum, 1986).
В классическом труде «Биосфера», опубликованном в 1926 г., В. И. Вернадский писал, что на земной поверхности нет химической силы, более постоянно действующей, а потому и более могущественной по своим конечным последствиям, чем живые организмы, взятые в целом. Следует отметить, что живые организмы необходимо рассматривать не как сумму автономных сил (в особенности трофических), а как системы, в которых активность организмов и популяций включена в определенные взаимодействующие и взаимосвязанные звенья.
Когда В. И. Вернадский создавал теорию биосферы, концепция трофических цепей базировалась на упрощенных и неполных представлениях, не позволяющих дать систематическое описание механизмов ассимиляции пищи, тем более в рамках единой науки - трофологии. Благодаря достижениям биологии последних лет представляется возможность охарактеризовать все процессы, происходящие в трофических цепях, на основании трех основных типов пищеварения, а также нескольких типов транспорта. Эти достижения позволяют также показать, что место вида в трофической цепи определяет его существование.
В сущности биология и физиология организмов тесно связаны с видовыми особенностями питания и добывания пищи. Экологи, и в частности Ю. Одум (Odum, 1986), подчеркивают, что главная побудительная причина активности всех животных - поиски нужной пищи в достаточном количестве. Твердо установлено, что структура и деятельность сообществ в значительной мере зависят от наличия пищи, а видовые и индивидуальные особенности многих процессов связаны с местом вида в трофической цепи и способом добывания пищи.
Парадоксальной на первый взгляд кажется другая связь: любой организм (точнее, вид) приспосабливается к тому, чтобы самому служить источником пищи (Уголев, 1980, 1986а). Для этого организм должен обладать такими свойствами, как определенная фагичность, т. е. доступность для другого организма в качестве источника пищи, а также трофичность, т.е. способность быть ассимилированным другим организмом. Некоторые рыбы, насекомые и растения, будучи вполне фагичными для многих организмов, из-за содержания в них токсических веществ не обладают трофичностью для ряда других живых существ.
На первый взгляд, такое положение трофологии может показаться и телеологичным, и противоречащим действительности, так как существует много данных в пользу того, что организмы вырабатывают специальные способы защиты от потенциального хищника.
Эта сторона проблемы детально освещена в прекрасном обзоре К. Шмидт-Ниельсена (Schmidt-Nielsen, 1982). Однако анализ показывает, что сочетание фагичности и трофичности обеспечивает процветание вида (разумеется, если оно не переходит определенных границ). Отсюда вытекают некоторые неожиданные следствия, в частности взаимная адаптация так называемых трофических партнеров (например, строгая взаимная зависимость свойств и численности популяций хищника и его жертвы). Так, в соответствии с гипотезой естественного равновесия (Hairstonetal., 1960), выеданию биомассы травоядными животными препятствуют хищники, ограничивающие численность этих травоядных. Только в таком случае возможна определенная стабильность популяции жертвы за счет того, что хищники будут питаться преимущественно ее больными, дефектными и стареющими членами. Совершенствование же жертвы может привести хищника к гибели от голода и вследствие этого к ухудшению популяции жертвы из-за отсутствия контроля со стороны хищника. Анализ свойств этой пары демонстрирует важную роль источника пищи и наличие обратной связи. Это было продемонстрировано в невольных экспериментах человека, когда, например, в каком-либо регионе полностью уничтожалось поголовье хищника, а в другом происходила искусственная интродукция хищника с помощью специальных мероприятий по охране этих животных.
Взаимное приспособление в последовательных парах хищник-жертва можно проиллюстрировать некоторыми примерами, поражающими сложностью и точностью адаптаций. В частности, Т. Торнтон охарактеризовал способ, каким хищники оберегают вид, которым они питаются. Морские звезды и офиуры имеют период физиологического голодания в 1-2 месяца, приуроченный к моменту оседания личинок пластиножаберных моллюсков - одного из основных пищевых объектов названных хищников. Личинки очень малы, и популяцию моллюсков морские звезды и офиуры могли бы истребить чрезвычайно быстро, но этого не происходит, так как хищники «теряют аппетит». Когда же за время голодания хищников масса личинок увеличивается на 2-3 порядка, у звезд и офиур наступает пора питания. Так с помощью периодического голодания хищники сохраняют источник своего питания (по: Уголев, 1980).
Примером взаимных адаптаций может служить эволюция взаимоотношений паразит-хозяин, при которой происходят уменьшение вредных для хозяина последствий паразитизма или инфекционного воздействия и переход к нейтральным или часто симбионтным взаимоотношениям.
Таким образом, необходимым условием процветания вида является его положение в трофической цепи. Это положение обеспечивается эффективностью взаимодействий не только с предшествующими, но и с последующими членами цепи. Другими словами, существенную роль играет не только источник питания и его эффективное поглощение, но и поедаемость данного члена цепи. В сущности механизмы и закономерности трофических взаимодействий являются одной из самых важных проблем трофологии.
В последние десятилетия накопились серьезные основания для утверждения, что в биоценозах представители различных групп организмов, в том числе далеких таксонов и даже различных царств, взаимодействуют не только с помощью массообмена, но и с помощью химических посредников. Примеры таких взаимодействий представлены в ряде блестящих обзоров (Rice, 1978; Сравнительная физиология..., 1978; Schmidt-Nielsen, 1982; Odum, 1986, и др.).
Регуляторные взаимодействия организмов, так называемая аллелопатия (термин предложил Н. Моlischв 1937 г.), на всех уровнях эволюционной лестницы, включая микроорганизмы, распространены очень широко. Таким образом, в первом приближении аллелопатия сводится к способности организмов определенных видов выделять физиологически активные вещества, действующие на организмы других видов. Классическим примером аллелопатии может служить образование антибиотиков и фитонцидов. Сейчас обнаружены многочисленные как токсические, так и стимулирующие вещества, вырабатываемые всеми живыми существами (см. также 9.7). Следовательно, состав любого биоценоза зависит не только от прямых трофических взаимодействий, но и от взаимодействий регуляторного характера.
1.6. Аутотрофность человечества
На пути превращения биосферы в ноосферу В. И. Вернадский (см. 1980) в качестве одного из важнейших условий выдвигал формирование социальной аутотрофности человека. Следует отметить, что в данном случае речь идет именно о социальной, а не о биологической аутотрофности. Последняя подразумевает превращение человека в организм, обладающий свойствами растений и бактерий синтезировать углеводы из углекислого газа при участии солнечной энергии и фиксировать атмосферный азот. Такое толкование безусловно неправильно и противоречит смыслу прогрессивной эволюции. (О прогрессивной эволюции см.: Завадский, Колчинский, 1977; Тимофеев-Ресовский и др., 1977; Эволюция, 1981; Развитие эволюционной теории..., 1983; Эволюционные идеи..., 1984; Проблемы..., 1985; Татаринов, 1987, и др.).
Действительно, по мнению многих эволюционистов, формирование двигательной активности, а в конечном итоге на ее основании нервной деятельности и мышления, стало возможным лишь в результате биотрофии. Именно биотрофия позволила отказаться от огромного количества сложных биохимических процессов и от необходимости постоянной связи человека с определенным локусом Земли (подробнее см. обзор: Уголев, 1985).
Таким образом, при сохранении и совершенствовании питания, характерного для человека, речь идет о том, чтобы человек не оказывал разрушительного воздействия на биосферу, которое с течением времени возрастает и приобретает катастрофический характер.
Обращая внимание на взаимоотношения человечества как части биосферы с биосферой в целом, В. И. Вернадский писал: «Человек неразрывно связан в одно целое с жизнью всех живых существ, существующих или когда-либо существовавших... Человек связан с этим целым еще благодаря питанию. Эта новая связь, как бы она ни была тесна и необходима, совсем иного порядка, чем непрестанное чередование поколений живых существ... Правда, что эта связь составляет часть великого геохимического явления в обороте химических элементов в биосфере, вызванного питанием организованных существ. Однако связь эта может быть изменена, не затронув стойкости жизненного процесса в целом» (Вернадский, 1980. С. 231).
В. И. Вернадский обращал также внимание на то, что современный тип питания, основанный на использовании сельскохозяйственных продуктов и продуктов животноводства, стал, с одной стороны, разрушительной силой глобального значения, с другой - не удовлетворяет потребностей человечества в пище. Выход В. И. Вернадский видит в создании социальной аутотрофности, т. е. в переходе к питанию синтетическими продуктами, обладающими всеми необходимыми для жизнедеятельности свойствами.
Эти идеи замечательны в ряде отношений. В частности, В. И. Вернадский во многом предвосхитил те мысли, которые развиваются и в наше время, например в рекомендациях ООН и ее комитетов. Эти организации полагают, что снабжение населения планеты пищей, положенное в основу одной из главных программ, является одной из четырех самых важных задач, стоящих перед нами. В. И. Вернадский прав, говоря о том, что традиционные методы хозяйствования ради удовлетворения пищевых потребностей населения приводят к тяжелейшим экологическим последствиям. Значение этих идей не только сохраняется в наше время, но и возрастает. Вместе с тем В. И. Вернадский развивал замечательную мысль П.-Э.-М. Бертло, что прямое (парентеральное) питание, т. е. введение полезных пищевых веществ непосредственно в кровь, преобразует человека в новое существо. Он оперировал, как и его предшественники, довольно простыми представлениями о питании, которые были в то время. По всей вероятности, проблема парентерального питания гораздо сложнее, и вопрос о том, каким образом будет происходить совершенствование человека, должен обсуждаться на основе новых данных науки, в особенности новых данных трофологии (см. гл. 3 и 5).
В своем дополнении к основному труду по аутотрофности человечества В. И. Вернадский отметил некоторые сложности, возникающие в связи с питанием людей и животных синтетической пищей. Анализ этих вопросов и экспериментальная проверка влияния синтетической элементной диеты на организм послужили одной из причин ревизии теории сбалансированного питания и формирования теории адекватного питания (см. гл. 3). Эта теория не отрицает возможности перехода от современного, так называемого сельскохозяйственного, питания человечества к социальной аутотрофности, т. е. к промышленному питанию. Она лишь указывает на необходимость того, чтобы технология приготовления пищи и свойства последней были адекватны естественным технологиям ассимиляции пищевых веществ организмами различных типов, в том числе человеком (см.: Уголев, 1987а).
Наконец, в рамках современных представлений о трофических связях следует иметь в виду не только проблему входа, т. е. снабжения человечества пищей, но и проблему выхода, т. е. разрушения пищевых отходов и продуктов метаболизма. Решение этих проблем позволит предохранить природную среду от повреждающих влиянии, связанных с питанием человека и животных. Эта проблема огромной важности приобретает все большее значение.
1.7. Классическая и естественная классификации организмов на основе трофических процессов
Ранее по типу питания все организмы в зависимости от источника потребляемого углерода подразделялись на две основные группы: аутотрофов, к которым относили большинство растений и некоторые бактерии, и гетеротрофов - всех животных. Принималось, что организмы первой из этих двух групп используют лишь неорганические вещества, а второй - органические вещества наряду с неорганическими. Высказано предположение разделить гетеротрофов на биофагов-организмов, потребляющих другие живые организмы, и сапрофагов-организмов, использующих мертвые органические остатки (Wiegert, Owen, 1971).
Однако деление организмов на аутотрофов и гетеротрофов не представляется вполне удачным. Строго говоря, полная аутотрофия существовать не может, так как все живые существа нуждаются в поступлении пищи извне, т. е. в экзотрофии. При этом используются как органические материалы, так и неорганические, а значит каждый организм в какой-то мере является гетеротрофом и не может быть полным аутотрофом в прямом смысле этого термина.
В связи с принципиальными дефектами существующих классификаций нами предложена новая естественная классификация организмов, которая охватывает все разнообразие типов питания (Уголев, 1980, 1985). На одном полюсе этой классификационной шкалы находятся полные абиотрофы, на другом - полные биотрофы, а вся шкала между этими крайними группами занята организмами с возрастающей долей биотрофии. Под абиотрофами понимаются организмы, питающиеся только неорганическими компонентами, под биотрофами - организмы, потребляющие в качестве пищи органические и неорганические вещества, содержащиеся в биологических объектах. Старое понятие «гетеротроф» может быть использовано для обобщенного описания частичных и полных биотрофов. Термин «аутотроф» кажется неоправданным и должен быть заменен термином «абиотроф» (см. также гл. 9).
Переходы от абиотрофии к биотрофии сложны и постепенны. В сущности полностью абиотрофных организмов, т. е. тех, которые синтезируют все необходимые органические компоненты из неорганических, сейчас фактически не существует. Например, фотосинтезирующие организмы нельзя считать полными абиотрофами, так как они обычно не способны к фиксации азота неорганического происхождения (азот минеральных солей, который используется растениями, на самом деле тоже образуется при разложении организмов). Следовательно, растения абиотрофны по большинству, но не по всем характеристикам. С другой стороны, азотфиксирующие бактерии абиотрофны по азоту, но для получения энергии используют биологические источники пищи. Таким образом, хотя полностью абиотрофных организмов почти не существует, есть абиотрофные системы, образуемые комплексом «растение-азотфиксирующие бактерии». В большинстве случаев предпочтительнее говорить не об абиотрофных организмах, а об абиотрофных сообществах, или комплексах. Эти комплексы включают в себя организмы, синтезирующие безазотистые органические вещества (углеводы, липиды и др.), но нуждающиеся в органическом азоте, который они получают от бактериальных сообществ, и абиотрофы, фиксирующие азот, но использующие органические источники углерода. Биотрофия у некоторых животных становится почти полной, например у хищников, однако они используют воду и соли небиологического происхождения. Полная биотрофия характерна для некоторых монофагов (организмов, питающихся единственным видом пищи), паразитов, эмбрионов, симбионтов и некоторых других.
Биотрофия в широком смысле существует в нескольких вариантах, имеющих различное биологическое значение, но близкие, а иногда и идентичные механизмы. Первый вариант - естественная экзотрофия, т. е. питание живыми организмами или их частями, второй - сапрофитизм, т. е. питание продуктами жизнедеятельности других организмов, отмирающими организмами, их частями. К биотрофии относится и эндотрофия - питание за счет внутренних депо и собственных структур тела или клетки, например темновое питание растений, а также ассимиляция собственного органического вещества голодающими животными и человеком.
Новая естественная классификация организмов на основе трофических процессов позволяет преодолеть еще некоторые трудности, хотя выводы подчас кажутся парадоксальными. Действительно, с этой точки зрения для растений углекислый газ служит экзо-нутриентом. Для азотфиксирующих бактерий такими нутриентами являются азот и водород (вода). Нетрудно видеть, что в этом случае обнаруживаются фундаментальные различия трофических процессов у растений и животных. Так, у растений существует дополнительный трофический ярус - преобразование первичных пищевых веществ, связанное с усвоением неорганических веществ и внешней энергии, во вторичные питательные вещества. В отличие от растений животные смогли прогрессировать, утратив многочисленные реакции, связанные с этим первым ярусом, причем у животных в большинстве случаев экзо- и эндонутриенты почти совпадают. Наконец, чрезвычайно важно, что в большинстве случаев обмен энергии в биологических объектах, использующих кислород, является замкнутым циклом, сходным с предложенным недавно техниками экологически чистым кислородноводородным технологическим циклом. Отличие заключается в том, что в большинстве случаев в биологических системах разделение водорода и кислорода не доводится до конца в том смысле, что выделения чистого водорода почти никогда не происходит. По-видимому, выделение чистого водорода было бы энергетически невыгодно, а кроме того, имело бы, по всей вероятности, глобальное отрицательное последствие - перемещение водорода в верхние слои атмосферы. В то же время включение водорода в соединения с углеродом дает возможность для построения экономичных запасов «топлива». Важно, что энергия составляет один из главных компонентов пищи, тогда как окислители не являются таковыми.
1.8. Происхождение и эволюция эндо- и экзотрофии.
Трофика и происхождение жизни
В свете современных знаний ясно, что механизмы эндотрофии и экзотрофии родственны, а не противоположны, как представлялось ранее, когда экзотрофию рассматривали в качестве гетеротрофии, а эндотрофию - в качестве аутотрофии. Становится понятным, например, структурное и функциональное сходство микроворсинок кишечника, обеспечивающего внешнюю биотрофию, и микроворсинок плаценты, реализующей питание зародыша за счет матери.
Однако вернемся к истокам жизни. Наиболее вероятно, что первичные носители жизни были примитивными и не имели сложного аппарата, необходимого для фиксации азота и фотосинтеза (см. также гл. 9). Они получали основные органические материалы в виде мономеров из небиологических источников (возможно, из омывающего их раствора). Следовательно, скорее всего, они были абиотрофами, потребляющими органические вещества. По-видимому, уже на ранних стадиях эволюции образовались ферментные системы, обеспечивающие частичное гидролитическое расщепление внутренних структур таких носителей жизни для использования их в качестве источника энергии и для построения новых структур. Такие гидролазы были, вероятно, наиболее древними. Можно предположить, что они первоначально обеспечивали перестройку структур и эндотрофию, а затем могли использоваться для утилизации соседних, но отмирающих организмов и их структур. Следовательно, гидролазы служили основой для формирования эндотрофии, а на более поздних этапах - экзотрофии.
Так, на базе первичной эндотрофии формировалась экзотрофия всех известных живых организмов. Эта древность происхождения и первичность эндотрофии позволяют понять сходство эндо- и экзотрофи- ческих процессов и осуществляющих их систем у столь далеких друг от друга организмов, как бактерии, высшие растения и животные. Кроме того, становится ясно, что все основные типы пищеварения сформировались на этой общей основе и сходны у всех организмов. Действительно, внеклеточное, мембранное и внутриклеточное пищеварение у всех живых организмов обладает многими общими чертами. Системы же фото- и хемосинтеза, необходимые для абиотрофии, - это более поздние и весьма специализированные достижения эволюции.
Принципиальное сходство механизмов ассимиляции пищевых веществ с помощью деполимеризующих систем (т. е. механизмов пищеварения) у большинства организмов имеет огромное адаптивное значение. Благодаря этому сходству организмы могут приспосабливаться к изменению места в трофической цепи (за исключением первого организма, у которого органические вещества синтезируются из неорганических). Другими словами, принципиально возможно превращение растительноядных организмов в хищников того или иного порядка или в паразитов; возможен переход от хищничества к сапрофитному питанию, и т. д.
На основе общих механизмов возникли такие специализированные способы питания, как эмбриональное и молочное. Таким образом, сходство, а иногда идентичность механизмов ассимиляции пищи у различных организмов - не случайность, а отражение эволюционной общности их происхождения.
Один из остро дискутируемых вопросов, возникающих при анализе происхождения жизни на Земле, - это вопрос о пищевых источниках для первичных, наиболее примитивных живых систем, не способных к сложным синтезам. Предполагается, что такие системы использовали первичный бульон, содержащий все необходимые мономеры (см. также гл. 9). Мы обсуждали вопрос о появлении наиболее древних форм гетеротрофии и высказали предположение, что они возникли на основе первичной абиотрофии, связанной с использованием собственных структур тела протобионтов под воздействием собственных гидролаз при нехватке пищевых ресурсов - протоголоде. Ферменты, осуществляющие гидролиз таких структур, могли затем использоваться для расщепления структур соседних протобионтов, особенно погибавших в силу тех или иных причин. Возможно, в таких случаях гидролиз происходил как под влиянием ферментов, выделяющихся поглощающим организмом (протосекреция), так и в результате аутолиза погибших протобионтов. Как отмечено выше, в ходе дальнейшей эволюции аутолиз, в частности индуцированный, получил большое распространение в животном мире в качестве механизма начальных стадий гидролиза пищи. В этой связи важны соображения Н. Горовица (Horowitz, 1945) о происхождении синтеза органических молекул и появлении аутотрофии. В 1945 г. он высказал предположение относительно того, что по мере исчерпания внешних пищевых ресурсов выживали лишь те формы протобионтов, которые были способны к синтезу недостающих пищевых веществ (см. гл. 9).
Следовательно, по всей вероятности, уже на ранних стадиях развития жизни трофические связи стали играть большую роль. При этом процесс эволюции в значительной степени определялся доступностью и качеством источников питания и энергии.
1.9. Замкнутые трофические системы
Решение многих задач на Земле и за ее пределами требует создания искусственных, полностью или почти полностью замкнутых трофических систем или даже небольших биосфер. В таких системах с участием организованных в трофические цепи организмов различных видов и должен происходить круговорот веществ, как правило, для поддержания жизни больших и малых сообществ людей или животных. Формирование искусственных замкнутых трофических систем и искусственных микробиосфер имеет непосредственное прикладное значение при освоении космического пространства, мирового океана и пр.
Проблема создания замкнутых трофических систем, в особенности необходимых при длительных космических полетах, давно волнует исследователей и мыслителей. По этому поводу были развиты многие фундаментальные идеи. В отношении таких конструируемых человеком систем были выдвинуты важные, хотя в ряде случаев и нереальные требования. Речь идет о том, что трофические системы должны быть в высокой степени продуктивными, надежными, должны обладать высокими скоростями и полнотой дезактивации токсических компонентов. Ясно, что реализовать такую систему исключительно трудно. Действительно, высказывались сомнения о возможности конструирования безопасной и надежной экосистемы (обзор: Odum, 1986). Тем не менее следует попытаться хотя бы определить максимальную емкость трофической системы, образно говоря, выяснить, каким должен быть маленький остров, пригодный для жизни Робинзона Крузо, если он будет накрыт прозрачным, но непроницаемым колпаком.
В качестве примера можно привести недавно разработанную модель искусственной биосферы (биосфера II), которая является стабильной замкнутой системой и необходима для жизни в различных областях космического пространства, в том числе на Луне и Марсе (обзор: Allen, Nelson,1986). Она должна моделировать условия жизни на Земле, для чего следует хорошо знать природные технологии нашей планеты. Кроме того, такая биосфера должна содержать инженерные, биологические, энергетические,нформационные открытые системы,живые системы, накапливающие свободную энергию, и т. д. Как и биосфера, искусственная биосфера должна включать в себя подлинную воду, воздух, скалы, землю, растительность и т. д. Она должна моделировать джунгли, пустыни, саванну, океан, болота, интенсивное земледелие и т. д., напоминающие родину человека (рис. 1.8). При этом оптимальное отношение искусственного океана и поверхности суши должно составлять не 70:30, как на Земле, а 15:85. Однако океан в искусственной биосфере должен быть по крайней мере в 10 раз более эффективным, чем настоящий.

| Рис. 1.8. Поперечный срез искусственной биосферы II (по: Allen, Nelson, 1986). |
Недавно эти же исследователи (Allen, Nelson, 1986) представили описание модельного комплекса связанных искусственных биосфер, разработанных для продолжительной жизни 64-80 человек на Марсе. Каждая из таких 4 биосфер, радиально расположенных по отношению к так называемому техническому центру, служит жизненным пространством для 6-10 человек. В техническом центре находится резервный океан для смягчения окружающей среды и поддержания замкнутой системы в целом. Существуют также биологическая, транспортная, горная и оперативная группы, а также госпиталь для визитеров с Земли, Луны или других частей Марса.
Конкретные проблемы питания в космосе при длительных полетах выходят за пределы этой книги. Тем не менее следует сказать, что при длительных полетах в космическом аппарате создается микромир, изолированный от привычной для человека среды на долгое, а в некоторых случаях и на неопределенно долгое время. Особенности этого микромира, и в частности особенности его трофики, во многом определяют существование системы в целом. По всей вероятности, одной из самых важных ступеней биотического круговорота служит деградация продуктов жизнедеятельности. Значение процессов деградации часто недооценивается. В частности, при обсуждении проблемы пищевых ресурсов человек традиционно рассматривается как высшее и конечное звено трофической цепи (обзоры: Odum, 1986; Biotechnology...,1989, и др.). Между тем такая постановка проблемы уже привела к формированию серьезных экологических дефектов, так как экологическая система может быть устойчивой лишь при сочетании эффективного поступления и расхода веществ. Примеры этому весьма многочисленны. К одному из них относится драматический эпизод в Австралии, где произошло разрушение растительных покровов пометом овец и коров из-за отсутствия жуков-навозников.
Во всех случаях проблемы деградации продуктов жизнедеятельности и элиминации самых ослабленных членов популяции чрезвычайно важны. Недавно развиваемая точка зрения неожиданно получила подтверждение. При моделировании длит
Дата добавления: 2016-06-22; просмотров: 1827;
Поиск по сайту
Узнать еще
- I. Демократия как тип политического режима.
- II. Агрессия как проявление побуждения
- II. КРОВЬ КАК ВНУТРЕННЯЯ СРЕДА ОРГАНИЗМА. ФИЗИОЛОГИЯ ЭРИТРОЦИТОВ И ЛЕЙКОЦИТОВ.
- IV. Глобальные проблемы современности.
- Windows 9X как пример системы, управляемой событиями
- Авангардизм как стадия развития искусства.
- Австралопитековые как переходное звено от высшей обезьяны к древнему человеку.
- Автоматизация как направление дальнейшего совершенствования управления силами и средствами

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
